

Cholecystokinin exerts an effect via the endocannabinoid system to inhibit GABAergic transmission in midbrain periaqueductal gray
- PMID: 21525858
- PMCID: PMC3154098
- DOI: 10.1038/npp.2011.59
Cholecystokinin exerts an effect via the endocannabinoid system to inhibit GABAergic transmission in midbrain periaqueductal gray
Abstract
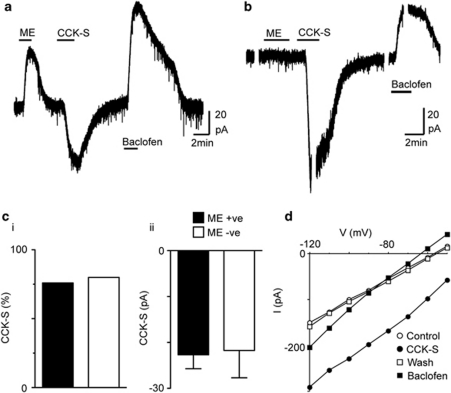
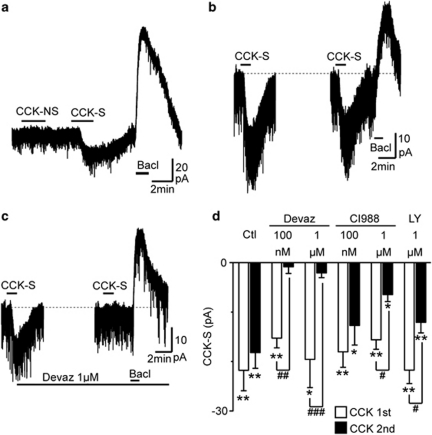
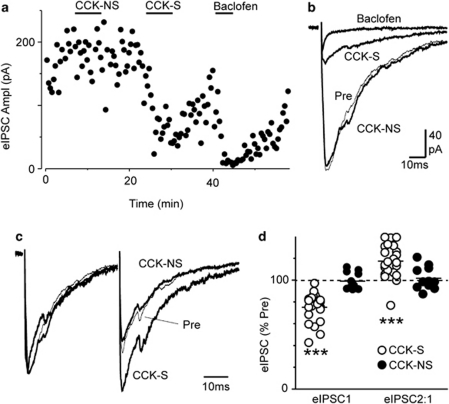
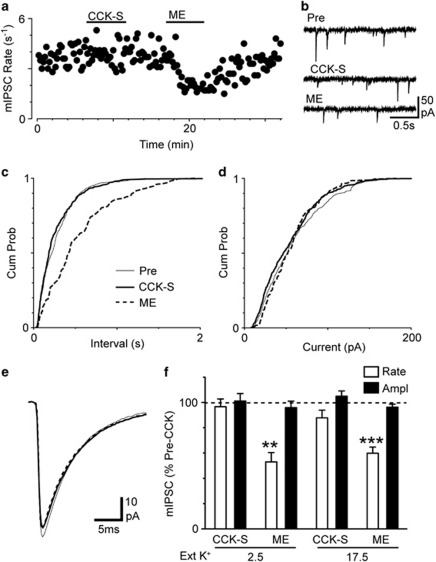
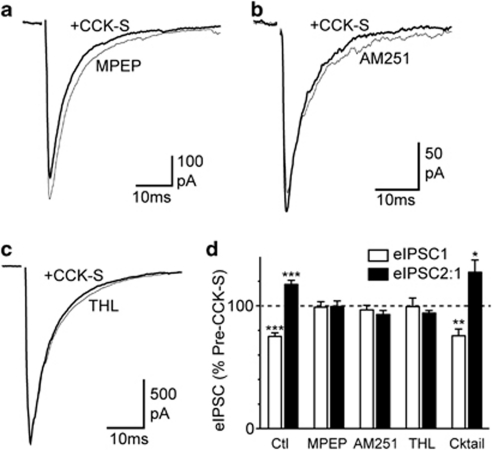
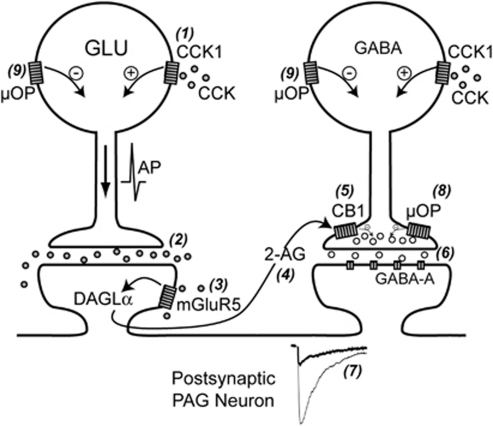
Cholecystokinin modulates pain and anxiety via its functions within brain regions such as the midbrain periaqueductal gray (PAG). The aim of this study was to examine the cellular actions of cholecystokinin on PAG neurons. Whole-cell patch clamp recordings were made from rat midbrain PAG slices in vitro to examine the postsynaptic effects of cholecystokinin and its effects on synaptic transmission. Sulfated cholecystokinin-(26-33) (CCK-S, 100-300 nM), but not non-sulfated cholecystokinin-(26-33) (CCK-NS, 100-300 nM) produced an inward current in a sub-population of opioid sensitive and insensitive PAG neurons, which did not reverse over a range of membrane potentials. The CCK-S-induced current was abolished by the CCK1 selective antagonist devazepide (100 nM), but not by the CCK2 selective antagonists CI988 (100 nM, 1 μM) and LY225910 (1 μM). CCK-S, but not CCK-NS produced a reduction in the amplitude of evoked GABA(A)-mediated inhibitory postsynaptic currents (IPSCs) and an increase in the evoked IPSC paired-pulse ratio. By contrast, CCK-S had little effect on the rate and amplitude of TTX-resistant miniature IPSCs under basal conditions and when external K(+) was elevated. The CCK-S-induced inhibition of evoked IPSCs was abolished by the cannabinoid CB1 receptor antagonist AM251 (3 μM), the mGluR5 antagonist MPEP (10 μM) and the 1, 2-diacylglycerol lipase (DAGLα) inhibitor tetrahydrolipstatin (10 μM). In addition, CCK-S produced an increase in the rate of spontaneous non-NMDA-mediated, TTX-dependent excitatory postsynaptic currents (EPSCs). These results suggest that cholecystokinin produces direct neuronal depolarisation via CCK1 receptors and inhibits GABAergic synaptic transmission via action potential-dependent release of glutamate and mGluR5-induced endocannabinoid signaling. Thus, cholecystokinin has cellular actions within the PAG that can both oppose and reinforce opioid and cannabinoid modulation of pain and anxiety within this brain structure.
Figures
References
-
- Bertoglio LJ, Zangrossi H., Jr Involvement of dorsolateral periaqueductal gray cholecystokinin-2 receptors in the regulation of a panic-related behavior in rats. Brain Res. 2005;1059:46–51. - PubMed
-
- Brack KE, Jeffery SM, Lovick TA. Cardiovascular and respiratory responses to a panicogenic agent in anaesthetised female Wistar rats at different stages of the oestrous cycle. Eur J Neurosci. 2006;23:3309–3318. - PubMed
-
- Brack KE, Lovick TA. Neuronal excitability in the periaqueductal grey matter during the estrous cycle in female Wistar rats. Neuroscience. 2007;144:325–335. - PubMed
-
- Carlberg M, Gundlach AL, Mercer LD, Beart PM. Autoradiographic localization of cholecystokinin A and B receptors in rat brain using [125I]d-Tyr25 (Nle28,31)-CCK 25–33S. Eur J Neurosci. 1992;4:563–573. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
