

Retinoic acid functions as a key GABAergic differentiation signal in the basal ganglia
- PMID: 21532733
- PMCID: PMC3075211
- DOI: 10.1371/journal.pbio.1000609
Retinoic acid functions as a key GABAergic differentiation signal in the basal ganglia
Abstract
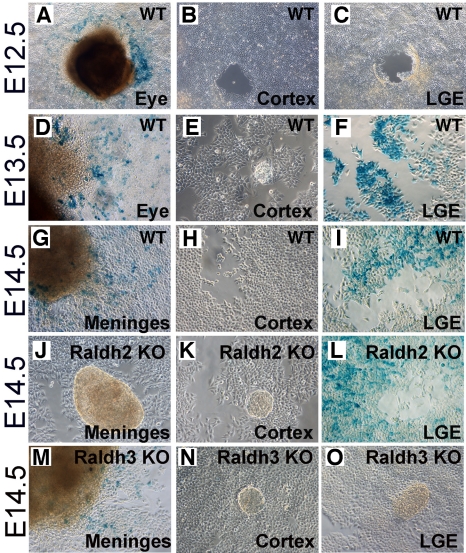
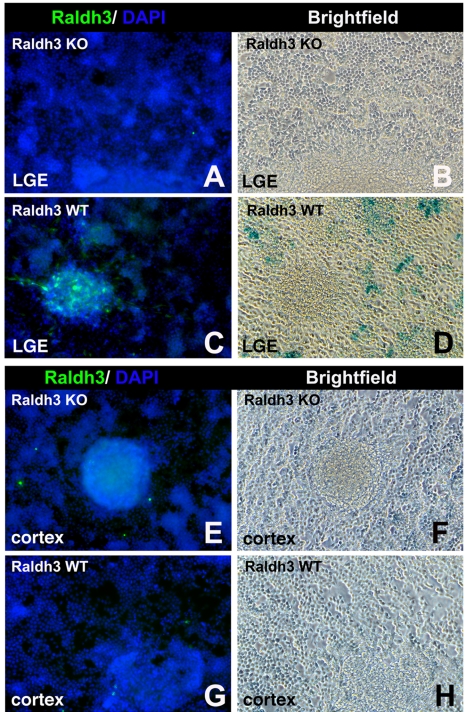
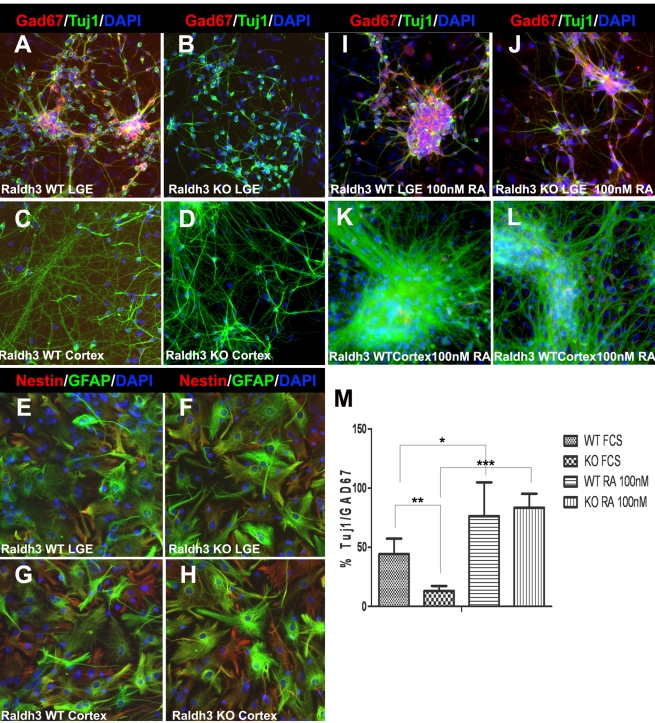
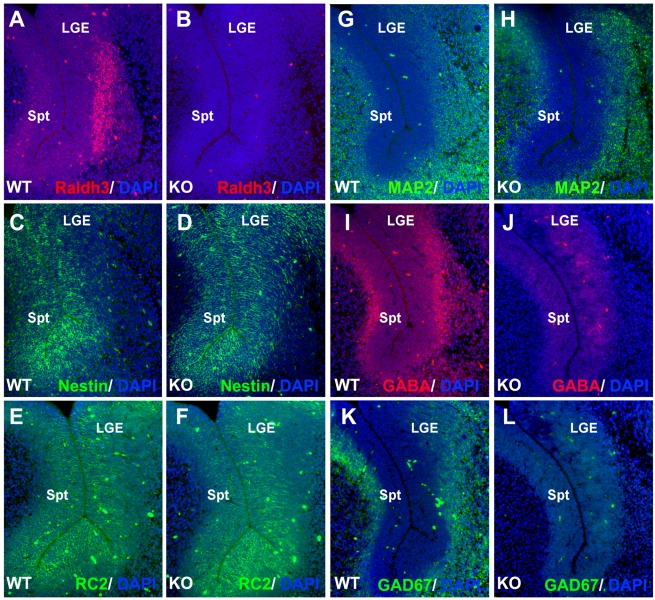
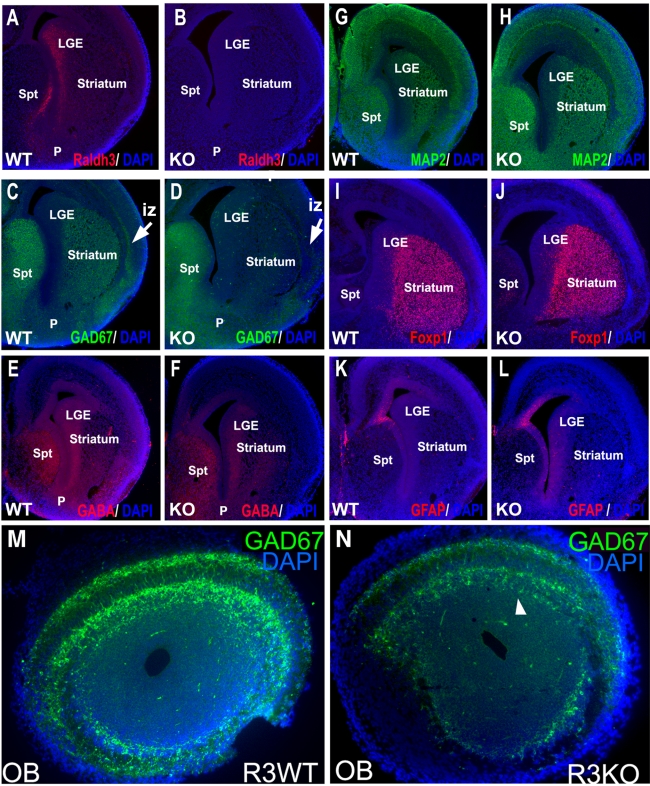
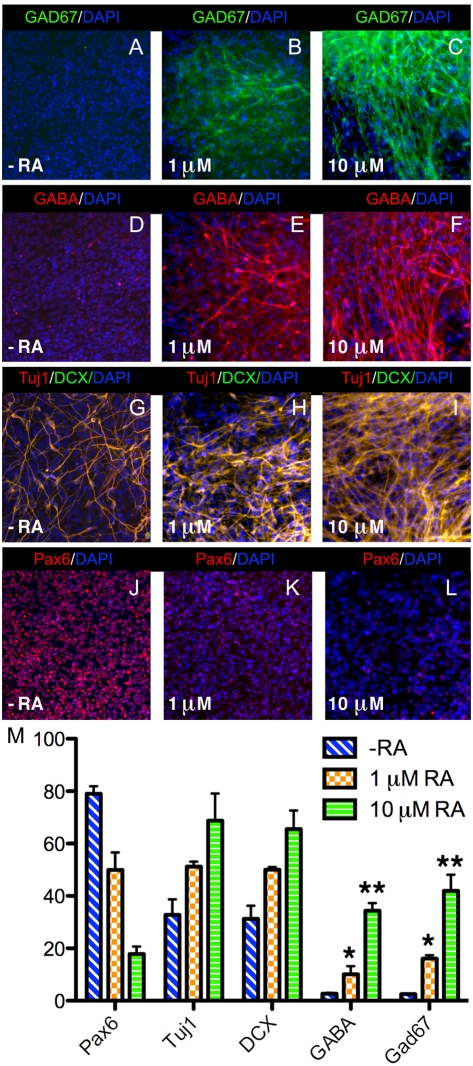
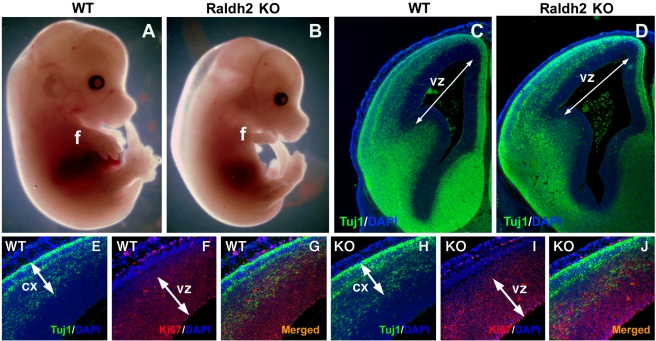
Although retinoic acid (RA) has been implicated as an extrinsic signal regulating forebrain neurogenesis, the processes regulated by RA signaling remain unclear. Here, analysis of retinaldehyde dehydrogenase mutant mouse embryos lacking RA synthesis demonstrates that RA generated by Raldh3 in the subventricular zone of the basal ganglia is required for GABAergic differentiation, whereas RA generated by Raldh2 in the meninges is unnecessary for development of the adjacent cortex. Neurospheres generated from the lateral ganglionic eminence (LGE), where Raldh3 is highly expressed, produce endogenous RA, which is required for differentiation to GABAergic neurons. In Raldh3⁻/⁻ embryos, LGE progenitors fail to differentiate into either GABAergic striatal projection neurons or GABAergic interneurons migrating to the olfactory bulb and cortex. We describe conditions for RA treatment of human embryonic stem cells that result in efficient differentiation to a heterogeneous population of GABAergic interneurons without the appearance of GABAergic striatal projection neurons, thus providing an in vitro method for generation of GABAergic interneurons for further study. Our observation that endogenous RA is required for generation of LGE-derived GABAergic neurons in the basal ganglia establishes a key role for RA signaling in development of the forebrain.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Moreno N, Gonzalez A, Retaux S. Development and evolution of the subpallium. Semin Cell Dev Biol. 2009;20:735–743. - PubMed
-
- Danglot L, Triller A, Marty S. The development of hippocampal interneurons in rodents. Hippocampus. 2006;16:1032–1060. - PubMed
-
- Rymar V. V, Sasseville R, Luk K. C, Sadikot A. F. Neurogenesis and stereological morphometry of calretinin-immunoreactive GABAergic interneurons of the neostriatum. J Comp Neurol. 2004;469:325–339. - PubMed
-
- Tepper J. M, Bolam J. P. Functional diversity and specificity of neostriatal interneurons. Curr Opin Neurobiol. 2004;14:685–692. - PubMed
-
- Anderson S. A, Marin O, Horn C, Jennings K, Rubenstein J. L. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development. 2001;128:353–363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
