

Analysis of the plant bos1 mutant highlights necrosis as an efficient defence mechanism during D. dadantii/Arabidospis thaliana interaction
- PMID: 21533045
- PMCID: PMC3080887
- DOI: 10.1371/journal.pone.0018991
Analysis of the plant bos1 mutant highlights necrosis as an efficient defence mechanism during D. dadantii/Arabidospis thaliana interaction
Abstract
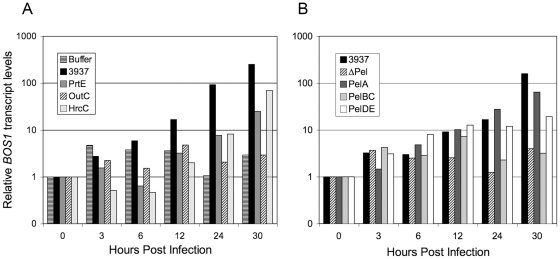
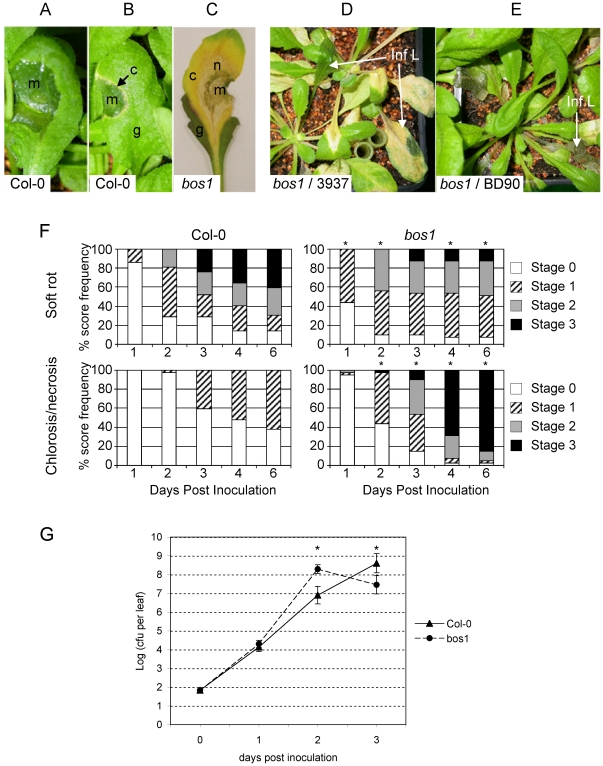
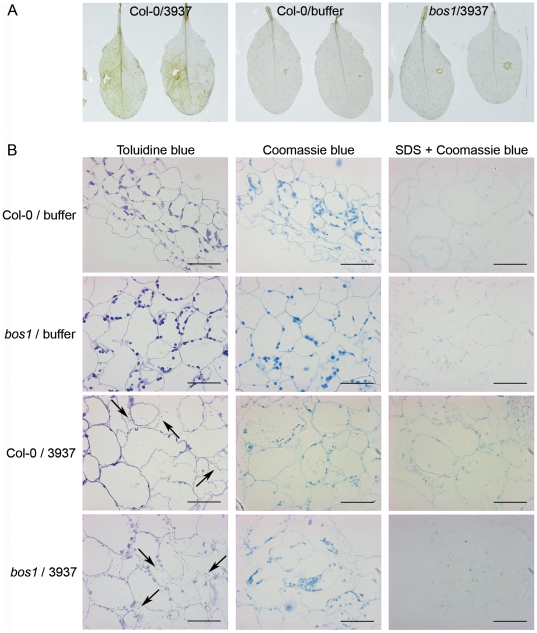
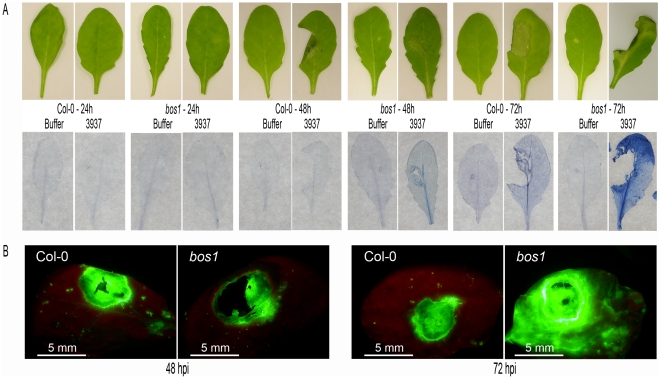
Dickeya dadantii is a broad host range phytopathogenic bacterium provoking soft rot disease on many plants including Arabidopsis. We showed that, after D. dadantii infection, the expression of the Arabidopsis BOS1 gene was specifically induced by the production of the bacterial PelB/C pectinases able to degrade pectin. This prompted us to analyze the interaction between the bos1 mutant and D. dadantii. The phenotype of the infected bos1 mutant is complex. Indeed, maceration symptoms occurred more rapidly in the bos1 mutant than in the wild type parent but at a later stage of infection, a necrosis developed around the inoculation site that provoked a halt in the progression of the maceration. This necrosis became systemic and spread throughout the whole plant, a phenotype reminiscent of that observed in some lesion mimic mutants. In accordance with the progression of maceration symptoms, bacterial population began to grow more rapidly in the bos1 mutant than in the wild type plant but, when necrosis appeared in the bos1 mutant, a reduction in bacterial population was observed. From the plant side, this complex interaction between D. dadantii and its host includes an early plant defence response that comprises reactive oxygen species (ROS) production accompanied by the reinforcement of the plant cell wall by protein cross-linking. At later timepoints, another plant defence is raised by the death of the plant cells surrounding the inoculation site. This plant cell death appears to constitute an efficient defence mechanism induced by D. dadantii during Arabidopsis infection.
Conflict of interest statement
Figures
References
-
- Perombelon MCM, Kelman A. Ecology of the soft rot erwinias. Annu Rev Phytopathol. 1980;18:361–387.
-
- Dellagi A, Rigault M, Segond D, Roux C, Kraepiel Y, et al. Siderophore-mediated upregulation of Arabidopsis ferritin expression in response to Erwinia chrysanthemi infection. Plant J. 2005;43:262–272. - PubMed
-
- Barras F, Van Gijsegem F, Chatterjee AK. Extracellular enzymes and pathogenesis of soft-rot Erwinia. Ann Rev Phytopathol. 1994;32:201–234.
-
- Collmer A, Keen N. The role of pectic enzymes in plant pathogenesis. Ann Rev Phytopathol. 1986;24:383–409.
-
- Kazemi-Pour N, Condemine G, Hugouvieux-Cotte-Pattat N. The secretome of the plant pathogenic bacterium Erwinia chrysanthemi. Proteomics. 2004;4:3177–3186. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
