

Hrk1 plays both Hog1-dependent and -independent roles in controlling stress response and antifungal drug resistance in Cryptococcus neoformans
- PMID: 21533251
- PMCID: PMC3076434
- DOI: 10.1371/journal.pone.0018769
Hrk1 plays both Hog1-dependent and -independent roles in controlling stress response and antifungal drug resistance in Cryptococcus neoformans
Abstract
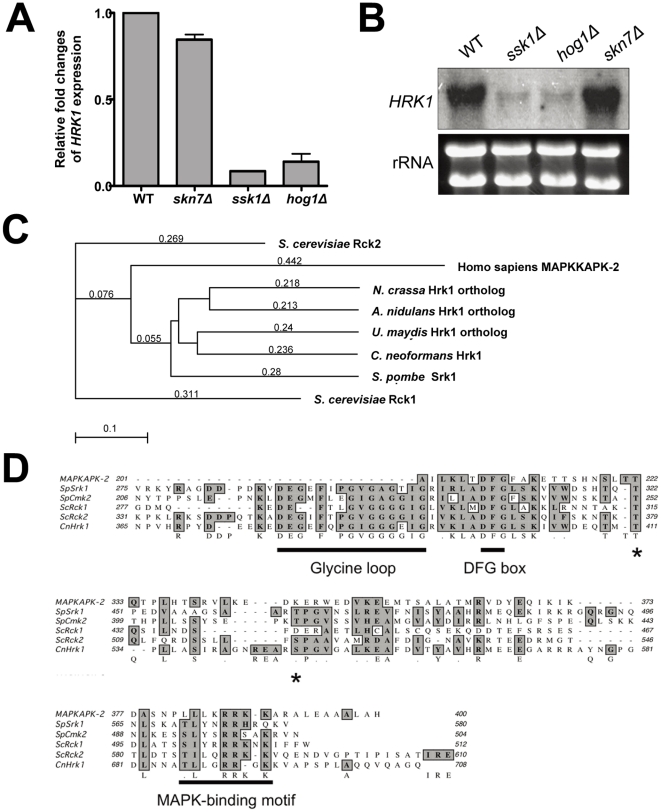
The HOG (High Osmolarity Glycerol response) pathway plays a central role in controlling stress response, ergosterol biosynthesis, virulence factor production, and differentiation of Cryptococcus neoformans, which causes fatal fungal meningoencephalitis. Recent transcriptome analysis of the HOG pathway discovered a Hog1-regulated gene (CNAG_00130.2), encoding a putative protein kinase orthologous to Rck1/2 in Saccharomyces cerevisiae and Srk1 in Schizosaccharomyces pombe. Its function is not known in C. neoformans. The present study functionally characterized the role of Hrk1 in C. neoformans. Northern blot analysis confirmed that HRK1 expression depends on the Hog1 MAPK. Similar to the hog1Δ mutant, the hrk1Δ mutant exhibited almost complete resistance to fludioxonil, which triggers glycerol biosynthesis via the HOG pathway. Supporting this, the hrk1Δ mutant showed reduced intracellular glycerol accumulation and swollen cell morphology in response to fludioxonil, further suggesting that Hrk1 works downstream of the HOG pathway. However, Hrk1 also appeared to have Hog1-independent functions. Mutation of HRK1 not only further increased osmosensitivity of the hog1Δ mutant, but also suppressed increased azole-resistance of the hog1Δ mutant in an Erg11-independent manner. Furthermore, unlike the hog1Δ mutant, Hrk1 was not involved in capsule biosynthesis. Hrk1 was slightly involved in melanin production but dispensable for virulence of C. neoformans. These findings suggest that Hrk1 plays both Hog1-dependent and -independent roles in stress and antifungal drug susceptibility and virulence factor production in C. neoformans. Particularly, the finding that inhibition of Hrk1 substantially increases azole drug susceptibility provides a novel strategy for combination antifungal therapy.
Conflict of interest statement
Figures







Similar articles
-
Calcineurin, Mpk1 and Hog1 MAPK pathways independently control fludioxonil antifungal sensitivity in Cryptococcus neoformans.Microbiology (Reading). 2006 Mar;152(Pt 3):591-604. doi: 10.1099/mic.0.28571-0. Microbiology (Reading). 2006. PMID: 16514140
-
A flucytosine-responsive Mbp1/Swi4-like protein, Mbs1, plays pleiotropic roles in antifungal drug resistance, stress response, and virulence of Cryptococcus neoformans.Eukaryot Cell. 2012 Jan;11(1):53-67. doi: 10.1128/EC.05236-11. Epub 2011 Nov 11. Eukaryot Cell. 2012. PMID: 22080454 Free PMC article.
-
Ssk2 mitogen-activated protein kinase kinase kinase governs divergent patterns of the stress-activated Hog1 signaling pathway in Cryptococcus neoformans.Eukaryot Cell. 2007 Dec;6(12):2278-89. doi: 10.1128/EC.00349-07. Epub 2007 Oct 19. Eukaryot Cell. 2007. PMID: 17951522 Free PMC article.
-
The Environmental Effects on Virulence Factors and the Antifungal Susceptibility of Cryptococcus neoformans.Int J Mol Sci. 2021 Jun 11;22(12):6302. doi: 10.3390/ijms22126302. Int J Mol Sci. 2021. PMID: 34208294 Free PMC article. Review.
-
Current understanding of HOG-MAPK pathway in Aspergillus fumigatus.Mycopathologia. 2013 Feb;175(1-2):13-23. doi: 10.1007/s11046-012-9600-5. Epub 2012 Nov 18. Mycopathologia. 2013. PMID: 23161019 Review.
Cited by
-
A Ferroxidase, Cfo1, Regulates Diverse Environmental Stress Responses of Cryptococcus neoformans through the HOG Pathway.Mycobiology. 2014 Jun;42(2):152-7. doi: 10.5941/MYCO.2014.42.2.152. Epub 2014 Jun 30. Mycobiology. 2014. PMID: 25071384 Free PMC article.
-
Glucan Unmasking Identifies Regulators of Temperature-Induced Translatome Reprogramming in C. neoformans.mSphere. 2021 Feb 10;6(1):e01281-20. doi: 10.1128/mSphere.01281-20. mSphere. 2021. PMID: 33568457 Free PMC article.
-
Identification of a major IP5 kinase in Cryptococcus neoformans confirms that PP-IP5/IP7, not IP6, is essential for virulence.Sci Rep. 2016 Apr 1;6:23927. doi: 10.1038/srep23927. Sci Rep. 2016. PMID: 27033523 Free PMC article.
-
The SrkA Kinase Is Part of the SakA Mitogen-Activated Protein Kinase Interactome and Regulates Stress Responses and Development in Aspergillus nidulans.Eukaryot Cell. 2015 May;14(5):495-510. doi: 10.1128/EC.00277-14. Epub 2015 Mar 27. Eukaryot Cell. 2015. PMID: 25820520 Free PMC article.
-
Targeting the oxidative stress response system of fungi with redox-potent chemosensitizing agents.Front Microbiol. 2012 Mar 16;3:88. doi: 10.3389/fmicb.2012.00088. eCollection 2012. Front Microbiol. 2012. PMID: 22438852 Free PMC article.
References
-
- Hohmann S, Krantz M, Nordlander B. Yeast osmoregulation. Methods Enzymol. 2007;428:29–45. - PubMed
-
- Barone FC, Irving EA, Ray AM, Lee JC, Kassis S, et al. Inhibition of p38 mitogen-activated protein kinase provides neuroprotection in cerebral focal ischemia. Med Res Rev. 2001;21:129–145. - PubMed
-
- Lee JC, Kumar S, Griswold DE, Underwood DC, Votta BJ, et al. Inhibition of p38 MAP kinase as a therapeutic strategy. Immunopharmacology. 2000;47:185–201. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
