

Iron-regulated lysis of recombinant Escherichia coli in host releases protective antigen and confers biological containment
- PMID: 21536797
- PMCID: PMC3191992
- DOI: 10.1128/IAI.01219-10
Iron-regulated lysis of recombinant Escherichia coli in host releases protective antigen and confers biological containment
Abstract
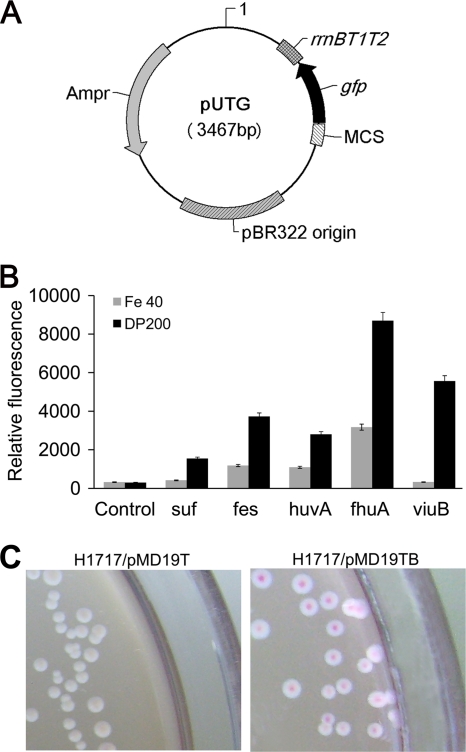
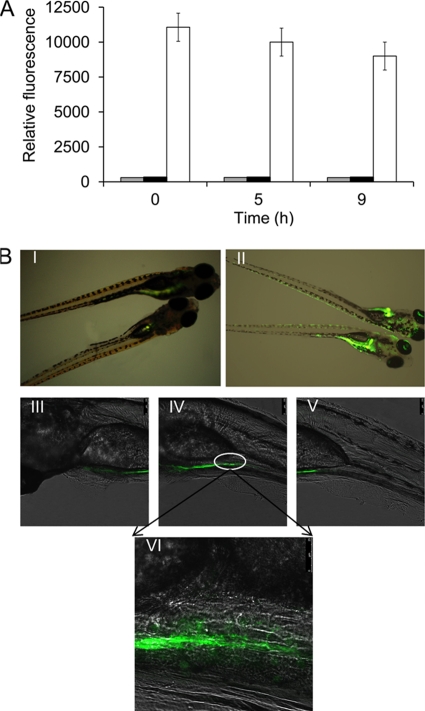
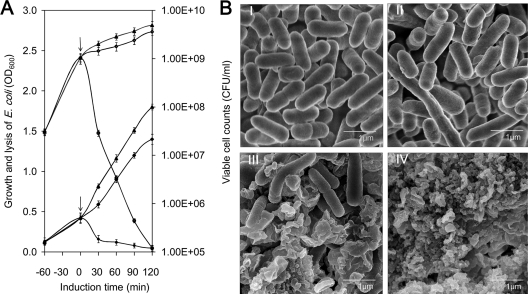
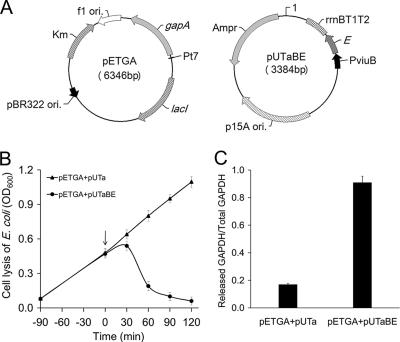
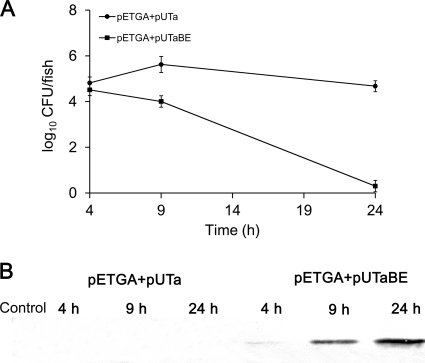
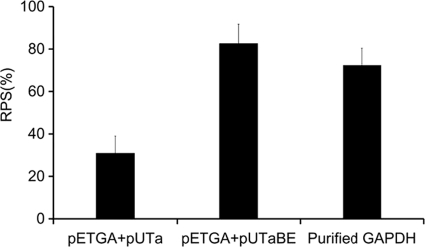
The use of a recombinant bacterial vector vaccine is an attractive vaccination strategy to induce an immune response to a carried protective antigen. The superiorities of live bacterial vectors include mimicry of a natural infection, intrinsic adjuvant properties, and the potential for administration by mucosal routes. Escherichia coli is a simple and efficient vector system for production of exogenous proteins. In addition, many strains are nonpathogenic and avirulent, making it a good candidate for use in recombinant vaccine design. In this study, we screened 23 different iron-regulated promoters in an E. coli BL21(DE3) vector and found one, P(viuB), with characteristics suitable for our use. We fused P(viuB) with lysis gene E, establishing an in vivo inducible lysis circuit. The resulting in vivo lysis circuit was introduced into a strain also carrying an IPTG (isopropyl-β-d-thiogalactopyranoside)-inducible P(T7)-controlled protein synthesis circuit, forming a novel E. coli-based protein delivery system. The recombinant E. coli produced a large amount of antigen in vitro and could deliver the antigen into zebrafish after vaccination via injection. The strain subsequently lysed in response to the iron-limiting signal in vivo, implementing antigen release and biological containment. The gapA gene, encoding the protective antigen GAPDH (glyceraldehyde-3-phosphate dehydrogenase) from the fish pathogen Aeromonas hydrophila LSA34, was introduced into the E. coli-based protein delivery system, and the resultant recombinant vector vaccine was evaluated in turbot (Scophtalmus maximus). Over 80% of the vaccinated fish survived challenge with A. hydrophila LSA34, suggesting that the E. coli-based antigen delivery system has great potential in bacterial vector vaccine applications.
Figures
Similar articles
-
A quorum sensing-based in vivo expression system and its application in multivalent bacterial vaccine.Microb Cell Fact. 2015 Mar 18;14:37. doi: 10.1186/s12934-015-0213-9. Microb Cell Fact. 2015. PMID: 25888727 Free PMC article.
-
Cell-penetrating peptides mediated protein cross-membrane delivery and its use in bacterial vector vaccine.Fish Shellfish Immunol. 2014 Jul;39(1):8-16. doi: 10.1016/j.fsi.2014.04.003. Epub 2014 Apr 16. Fish Shellfish Immunol. 2014. PMID: 24746937
-
A novel in vivo inducible expression system in Edwardsiella tarda for potential application in bacterial polyvalence vaccine.Fish Shellfish Immunol. 2011 Dec;31(6):1097-105. doi: 10.1016/j.fsi.2011.09.015. Epub 2011 Sep 25. Fish Shellfish Immunol. 2011. PMID: 21964456
-
A stable plasmid system for heterologous antigen expression in attenuated Vibrio anguillarum.Vaccine. 2011 Sep 16;29(40):6986-93. doi: 10.1016/j.vaccine.2011.07.036. Epub 2011 Jul 23. Vaccine. 2011. PMID: 21791231
-
Lactobacillus Mucosal Vaccine Vectors: Immune Responses against Bacterial and Viral Antigens.mSphere. 2018 May 16;3(3):e00061-18. doi: 10.1128/mSphere.00061-18. eCollection 2018 May-Jun. mSphere. 2018. PMID: 29769376 Free PMC article. Review.
Cited by
-
Development of a Fur-dependent and tightly regulated expression system in Escherichia coli for toxic protein synthesis.BMC Biotechnol. 2013 Mar 19;13:25. doi: 10.1186/1472-6750-13-25. BMC Biotechnol. 2013. PMID: 23510048 Free PMC article.
-
In Vivo Programmed Gene Expression Based on Artificial Quorum Networks.Appl Environ Microbiol. 2015 Aug;81(15):4984-92. doi: 10.1128/AEM.01113-15. Epub 2015 May 15. Appl Environ Microbiol. 2015. PMID: 25979894 Free PMC article.
-
Live-Attenuated Bacterial Vectors: Tools for Vaccine and Therapeutic Agent Delivery.Vaccines (Basel). 2015 Nov 10;3(4):940-72. doi: 10.3390/vaccines3040940. Vaccines (Basel). 2015. PMID: 26569321 Free PMC article. Review.
-
A quorum sensing-based in vivo expression system and its application in multivalent bacterial vaccine.Microb Cell Fact. 2015 Mar 18;14:37. doi: 10.1186/s12934-015-0213-9. Microb Cell Fact. 2015. PMID: 25888727 Free PMC article.
References
-
- Andrews S., Robinson A., Rodríguez-Quiones F. 2003. Bacterial iron homeostasis. FEMS Microbiol. Rev. 27:215–237 - PubMed
-
- Barry E., Altboum Z., Losonsky G., Levine M. 2003. Immune responses elicited against multiple enterotoxigenic Escherichia coli fimbriae and mutant LT expressed in attenuated Shigella vaccine strains. Vaccine 21:333–340 - PubMed
-
- Braun V. 2003. Iron uptake by Escherichia coli. Front. Biosci. 8:1409–1421 - PubMed
-
- Bury N., Grosell M. 2003. Waterborne iron acquisition by a freshwater teleost fish, zebrafish Danio rerio. J. Exp. Biol. 206:3529–3535 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
