

Astroglial networks scale synaptic activity and plasticity
- PMID: 21536893
- PMCID: PMC3100942
- DOI: 10.1073/pnas.1016650108
Astroglial networks scale synaptic activity and plasticity
Abstract
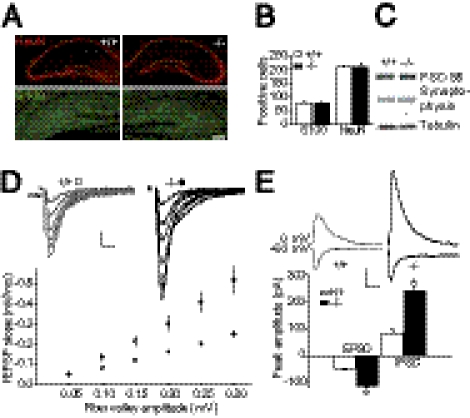
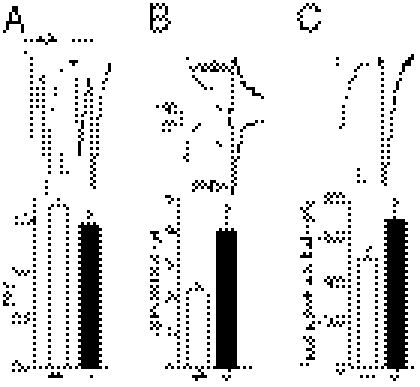
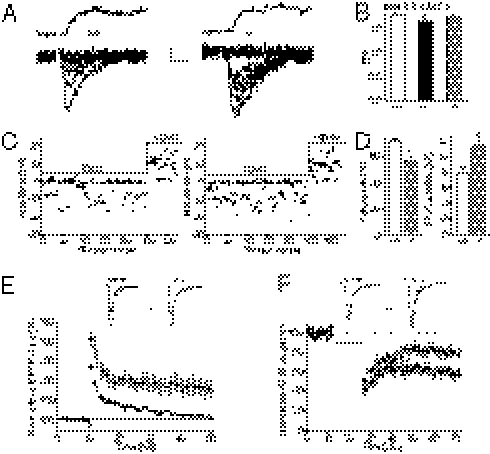
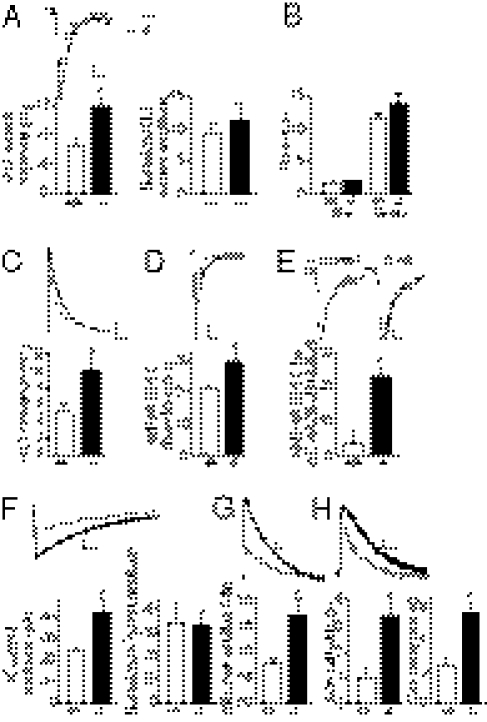
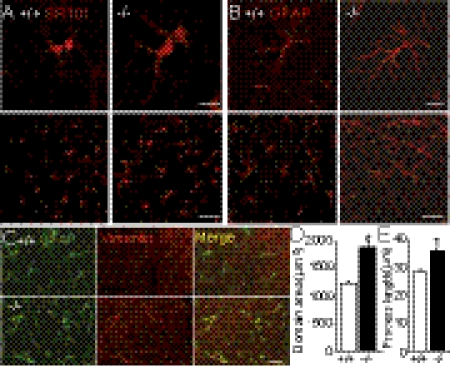
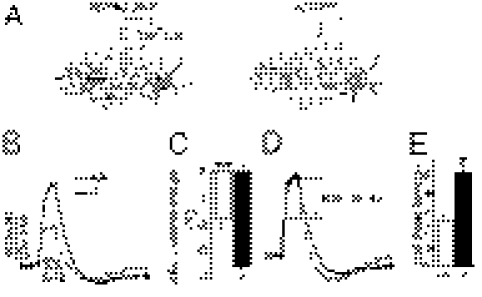
Astrocytes dynamically interact with neurons to regulate synaptic transmission. Although the gap junction proteins connexin 30 (Cx30) and connexin 43 (Cx43) mediate the extensive network organization of astrocytes, their role in synaptic physiology is unknown. Here we show, by inactivating Cx30 and Cx43 genes, that astroglial networks tone down hippocampal synaptic transmission in CA1 pyramidal neurons. Gap junctional networking facilitates extracellular glutamate and potassium removal during synaptic activity through modulation of astroglial clearance rate and extracellular space volume. This regulation limits neuronal excitability, release probability, and insertion of postsynaptic AMPA receptors, silencing synapses. By controlling synaptic strength, connexins play an important role in synaptic plasticity. Altogether, these results establish connexins as critical proteins for extracellular homeostasis, important for the formation of functional synapses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Perea G, Navarrete M, Araque A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009;32:421–431. - PubMed
-
- Giaume C, Koulakoff A, Roux L, Holcman D, Rouach N. Astroglial networks: A step further in neuroglial and gliovascular interactions. Nat Rev Neurosci. 2010;11:87–99. - PubMed
-
- Rouach N, Koulakoff A, Abudara V, Willecke K, Giaume C. Astroglial metabolic networks sustain hippocampal synaptic transmission. Science. 2008;322:1551–1555. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
