

Deficient serotonin neurotransmission and depression-like serotonin biomarker alterations in tryptophan hydroxylase 2 (Tph2) loss-of-function mice
- PMID: 21537332
- PMCID: PMC3536482
- DOI: 10.1038/mp.2011.50
Deficient serotonin neurotransmission and depression-like serotonin biomarker alterations in tryptophan hydroxylase 2 (Tph2) loss-of-function mice
Abstract
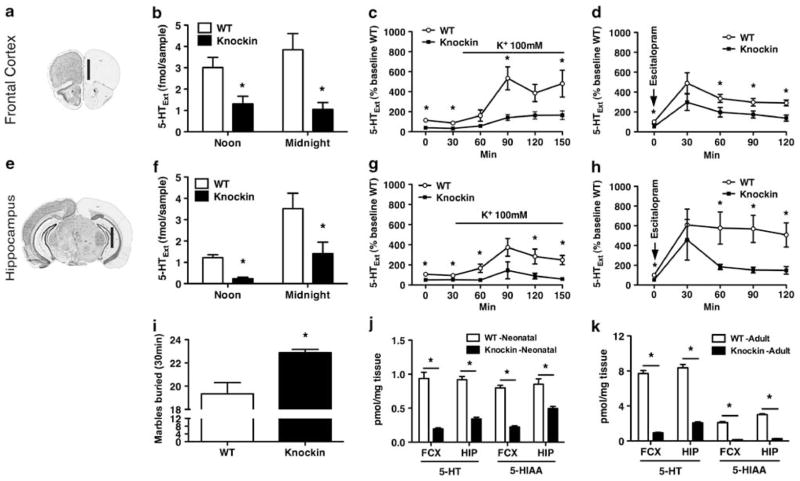
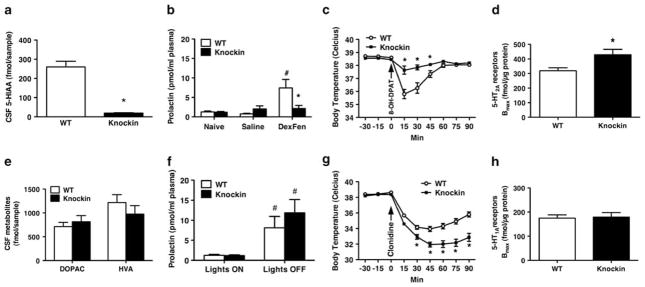
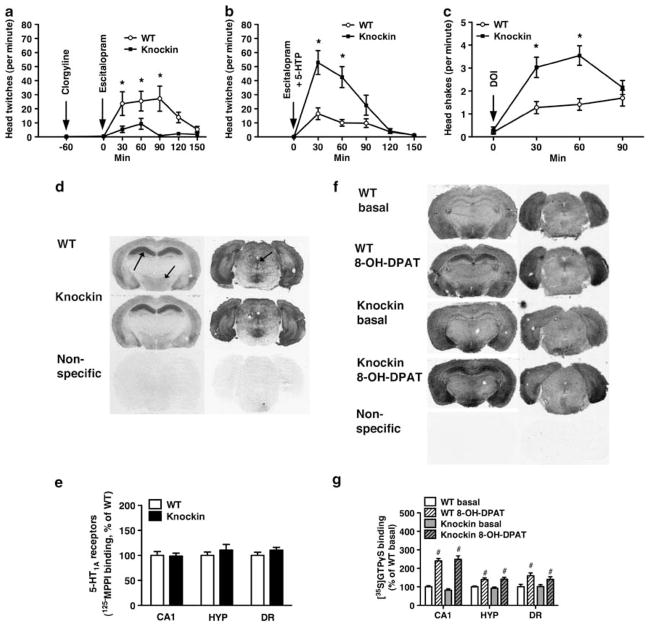
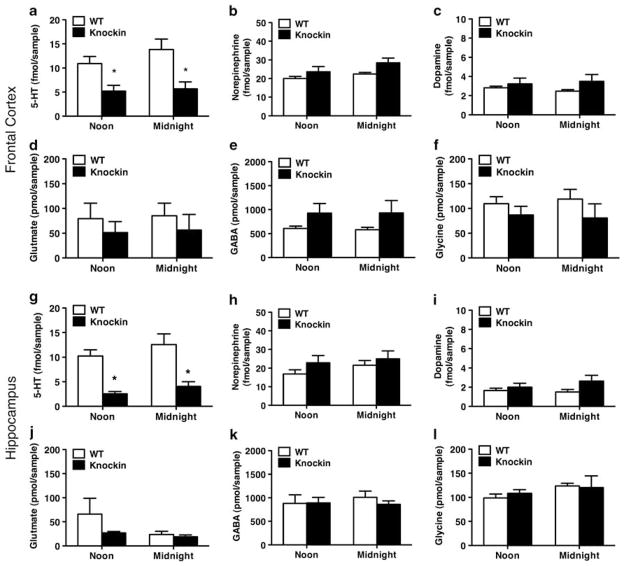
Probably the foremost hypothesis of depression is the 5-hydroxytryptamine (5-HT, serotonin) deficiency hypothesis. Accordingly, anomalies in putative 5-HT biomarkers have repeatedly been reported in depression patients. However, whether such anomalies in fact reflect deficient central 5-HT neurotransmission remains unresolved. We employed a naturalistic model of 5-HT deficiency, the tryptophan hydroxylase 2 (Tph2) R439H knockin mouse, to address this question. We report that Tph2 knockin mice have reduced basal and stimulated levels of extracellular 5-HT (5-HT(Ext)). Interestingly, cerebrospinal fluid (CSF) 5-hydroxyindoleacetic acid (5-HIAA) and fenfluramine-induced plasma prolactin levels are markedly diminished in the Tph2 knockin mice. These data seemingly confirm that low CSF 5-HIAA and fenfluramine-induced plasma prolactin reflects chronic, endogenous central nervous system (CNS) 5-HT deficiency. Moreover, 5-HT(1A) receptor agonist-induced hypothermia is blunted and frontal cortex 5-HT(2A) receptors are increased in the Tph2 knockin mice. These data likewise parallel core findings in depression, but are usually attributed to anomalies in the respective receptors rather than resulting from CNS 5-HT deficiency. Further, 5-HT(2A) receptor function is enhanced in the Tph2 knockin mice. In contrast, 5-HT(1A) receptor levels and G-protein coupling is normal in Tph2 knockin mice, indicating that the blunted hypothermic response relates directly to the low 5-HT(Ext). Thus, we show that not only low CSF 5-HIAA and a blunted fenfluramine-induced prolactin response, but also blunted 5-HT(1A) agonist-induced hypothermia and increased 5-HT(2A) receptor levels are bona fide biomarkers of chronic, endogenous 5-HT deficiency. Potentially, some of these biomarkers could identify patients likely to have 5-HT deficiency. This could have clinical research utility or even guide pharmacotherapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Coppen A. The biochemistry of affective disorders. Br J Psychiatry. 1967;113:1237–1264. - PubMed
-
- Shaw DM, Camps FE, Eccleston EG. 5-Hydroxytryptamine in the hind-brain of depressive suicides. Br J Psychiatry. 1967;113:1407–1411. - PubMed
-
- Meltzer H. Serotonergic dysfunction in depression. Br J Psychiatry Suppl. 1989;8:25–31. - PubMed
-
- Asberg M. Neurotransmitters and suicidal behavior. The evidence from cerebrospinal fluid studies. Ann NY Acad Sci. 1997;836:158–181. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
