

Down-regulation of hepatic urea synthesis by oxypurines: xanthine and uric acid inhibit N-acetylglutamate synthase
- PMID: 21540182
- PMCID: PMC3121350
- DOI: 10.1074/jbc.M110.209023
Down-regulation of hepatic urea synthesis by oxypurines: xanthine and uric acid inhibit N-acetylglutamate synthase
Abstract
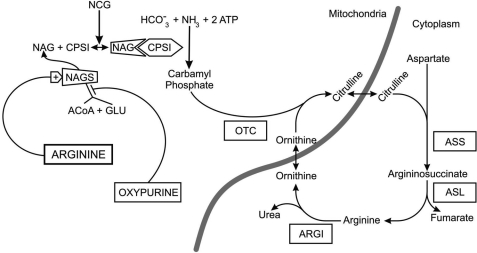
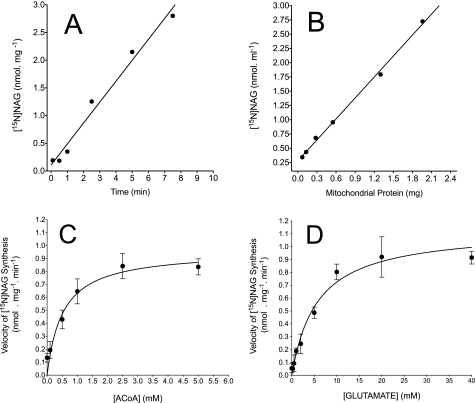
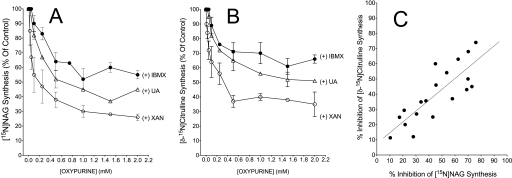
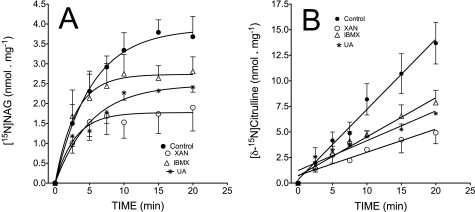
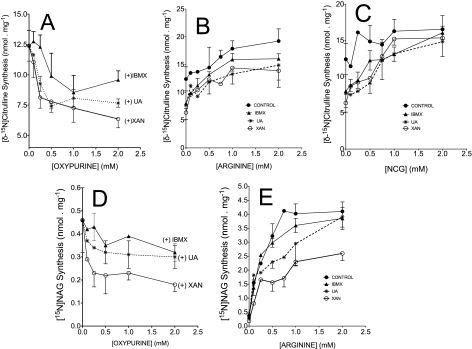
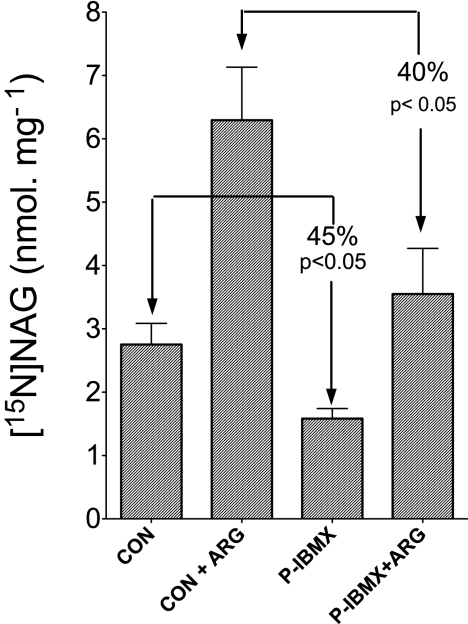
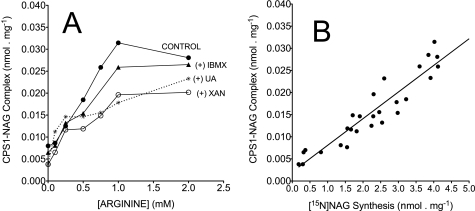
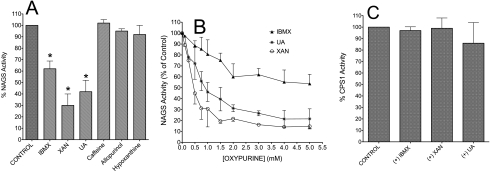
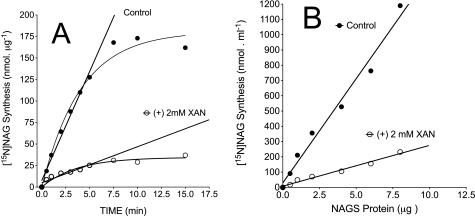
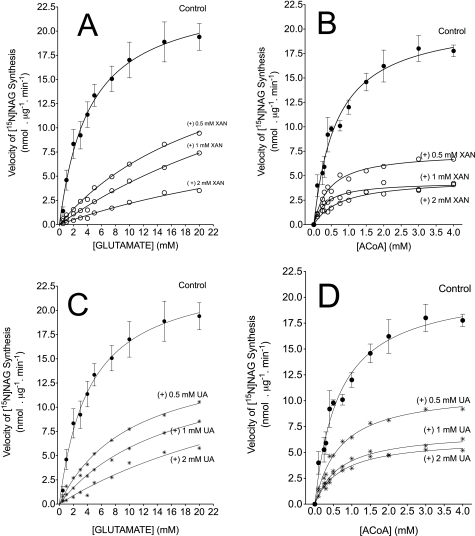
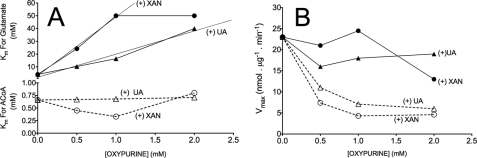
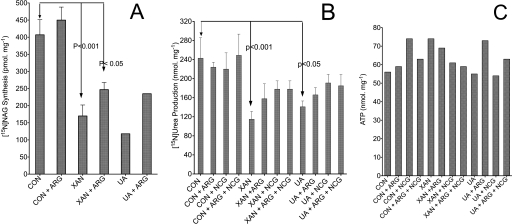
We previously reported that isobutylmethylxanthine (IBMX), a derivative of oxypurine, inhibits citrulline synthesis by an as yet unknown mechanism. Here, we demonstrate that IBMX and other oxypurines containing a 2,6-dione group interfere with the binding of glutamate to the active site of N-acetylglutamate synthetase (NAGS), thereby decreasing synthesis of N-acetylglutamate, the obligatory activator of carbamoyl phosphate synthase-1 (CPS1). The result is reduction of citrulline and urea synthesis. Experiments were performed with (15)N-labeled substrates, purified hepatic CPS1, and recombinant mouse NAGS as well as isolated mitochondria. We also used isolated hepatocytes to examine the action of various oxypurines on ureagenesis and to assess the ameliorating affect of N-carbamylglutamate and/or l-arginine on NAGS inhibition. Among various oxypurines tested, only IBMX, xanthine, or uric acid significantly increased the apparent K(m) for glutamate and decreased velocity of NAGS, with little effect on CPS1. The inhibition of NAGS is time- and dose-dependent and leads to decreased formation of the CPS1-N-acetylglutamate complex and consequent inhibition of citrulline and urea synthesis. However, such inhibition was reversed by supplementation with N-carbamylglutamate. The data demonstrate that xanthine and uric acid, both physiologically occurring oxypurines, inhibit the hepatic synthesis of N-acetylglutamate. An important and novel concept emerging from this study is that xanthine and/or uric acid may have a role in the regulation of ureagenesis and, thus, nitrogen homeostasis in normal and disease states.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK064913/DK/NIDDK NIH HHS/United States
- DK-053761/DK/NIDDK NIH HHS/United States
- HD058567/HD/NICHD NIH HHS/United States
- P30 HD026979/HD/NICHD NIH HHS/United States
- R56 DK053761/DK/NIDDK NIH HHS/United States
- R01 DK047870/DK/NIDDK NIH HHS/United States
- U54RR019453/RR/NCRR NIH HHS/United States
- R01 DK053761/DK/NIDDK NIH HHS/United States
- R01 HD058567/HD/NICHD NIH HHS/United States
- DK-R56-053761/DK/NIDDK NIH HHS/United States
- R01DK047870/DK/NIDDK NIH HHS/United States
- P01HD26979/HD/NICHD NIH HHS/United States
- R01DK064913/DK/NIDDK NIH HHS/United States
- U54 RR019453/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
