

Two distinct conformations of helix 6 observed in antagonist-bound structures of a beta1-adrenergic receptor
- PMID: 21540331
- PMCID: PMC3100933
- DOI: 10.1073/pnas.1100185108
Two distinct conformations of helix 6 observed in antagonist-bound structures of a beta1-adrenergic receptor
Abstract
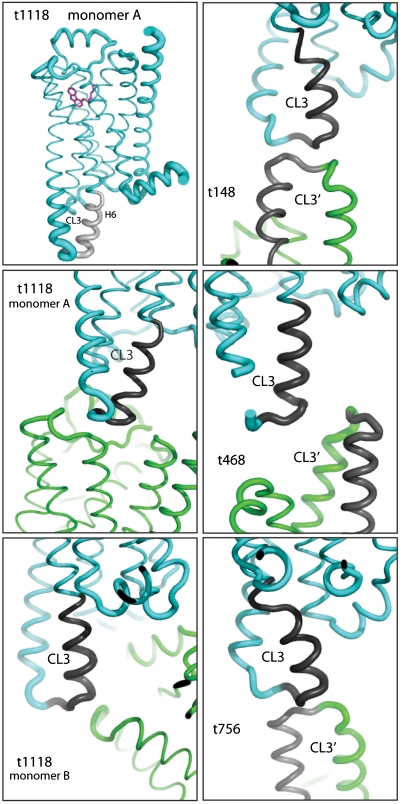
The β(1)-adrenergic receptor (β(1)AR) is a G-protein-coupled receptor whose inactive state structure was determined using a thermostabilized mutant (β(1)AR-M23). However, it was not thought to be in a fully inactivated state because there was no salt bridge between Arg139 and Glu285 linking the cytoplasmic ends of transmembrane helices 3 and 6 (the R(3.50) - D/E(6.30) "ionic lock"). Here we compare eight new structures of β(1)AR-M23, determined from crystallographically independent molecules in four different crystals with three different antagonists bound. These structures are all in the inactive R state and show clear electron density for cytoplasmic loop 3 linking transmembrane helices 5 and 6 that had not been seen previously. Despite significantly different crystal packing interactions, there are only two distinct conformations of the cytoplasmic end of helix 6, bent and straight. In the bent conformation, the Arg139-Glu285 salt bridge is present, as in the crystal structure of dark-state rhodopsin. The straight conformation, observed in previously solved structures of β-receptors, results in the ends of helices 3 and 6 being too far apart for the ionic lock to form. In the bent conformation, the R(3.50)-E(6.30) distance is significantly longer than in rhodopsin, suggesting that the interaction is also weaker, which could explain the high basal activity in β(1)AR compared to rhodopsin. Many mutations that increase the constitutive activity of G-protein-coupled receptors are found in the bent region at the cytoplasmic end of helix 6, supporting the idea that this region plays an important role in receptor activation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Fredriksson R, Schioth HB. The repertoire of G-protein-coupled receptors in fully sequenced genomes. Mol Pharmacol. 2005;67:1414–1425. - PubMed
-
- Foord SM, et al. International Union of Pharmacology. XLVI. G protein-coupled receptor list. Pharmacol Rev. 2005;57:279–288. - PubMed
-
- Li J, Edwards PC, Burghammer M, Villa C, Schertler GF. Structure of bovine rhodopsin in a trigonal crystal form. J Mol Biol. 2004;343:1409–1438. - PubMed
-
- Okada T, et al. The retinal conformation and its environment in rhodopsin in light of a new 2.2 Å crystal structure. J Mol Biol. 2004;342:571–583. - PubMed
-
- Palczewski K, et al. Crystal structure of rhodopsin: A G protein-coupled receptor. Science. 2000;289:739–745. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
