

Protective T cell immunity in mice following protein-TLR7/8 agonist-conjugate immunization requires aggregation, type I IFN, and multiple DC subsets
- PMID: 21540549
- PMCID: PMC3083762
- DOI: 10.1172/JCI45416
Protective T cell immunity in mice following protein-TLR7/8 agonist-conjugate immunization requires aggregation, type I IFN, and multiple DC subsets
Abstract
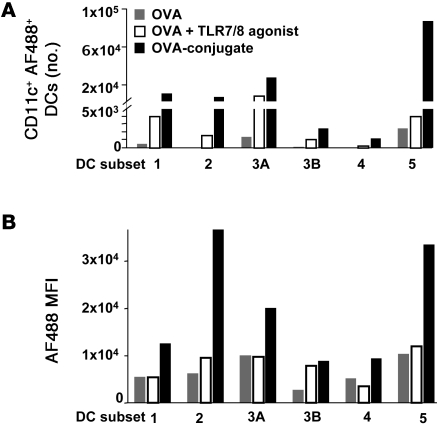
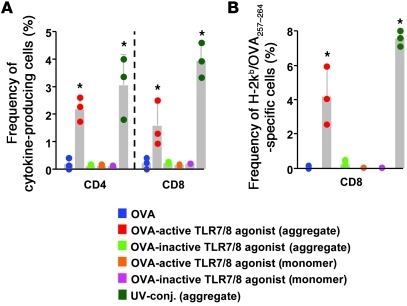
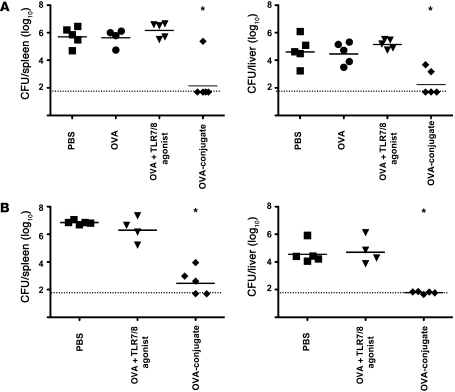
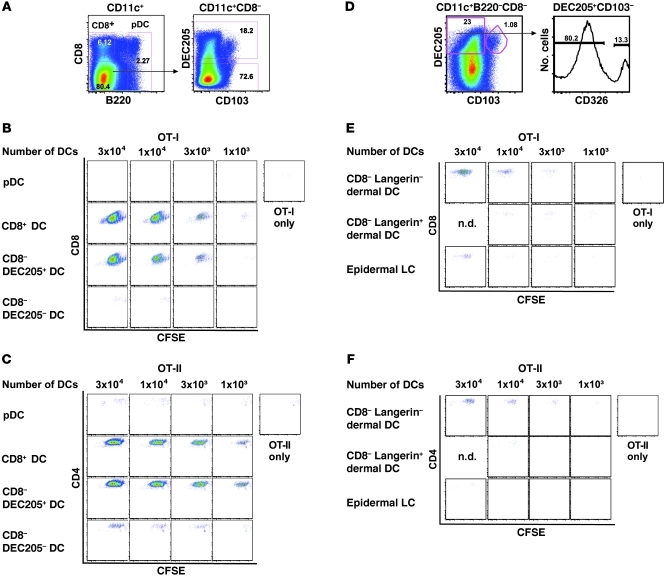
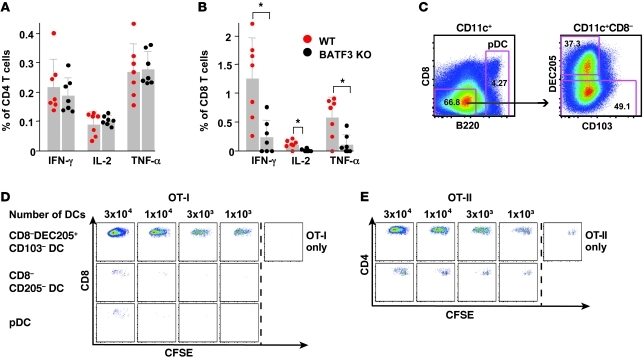
The success of a non-live vaccine requires improved formulation and adjuvant selection to generate robust T cell immunity following immunization. Here, using protein linked to a TLR7/8 agonist (conjugate vaccine), we investigated the functional properties of vaccine formulation, the cytokines, and the DC subsets required to induce protective multifunctional T cell immunity in vivo. The conjugate vaccine required aggregation of the protein to elicit potent Th1 CD4+ and CD8+ T cell responses. Remarkably, the conjugate vaccine, through aggregation of the protein and activation of TLR7 in vivo, led to an influx of migratory DCs to the LN and increased antigen uptake by several resident and migratory DC subsets, with the latter effect strongly influenced by vaccine-induced type I IFN. Ex vivo migratory CD8-DEC205+CD103-CD326- langerin-negative dermal DCs were as potent in cross-presenting antigen to naive CD8+ T cells as CD11c+CD8+ DCs. Moreover, these cells also influenced Th1 CD4+ T cell priming. In summary, we propose a model in which broad-based T cell-mediated responses upon vaccination can be maximized by codelivery of aggregated protein and TLR7/8 agonist, which together promote optimal antigen acquisition and presentation by multiple DC subsets in the context of critical proinflammatory cytokines.
Figures



References
-
- Trinchieri G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat Rev Immunol. 2003;3(2):133–146. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
