

Rostral growth of commissural axons requires the cell adhesion molecule MDGA2
- PMID: 21542908
- PMCID: PMC3113314
- DOI: 10.1186/1749-8104-6-22
Rostral growth of commissural axons requires the cell adhesion molecule MDGA2
Abstract
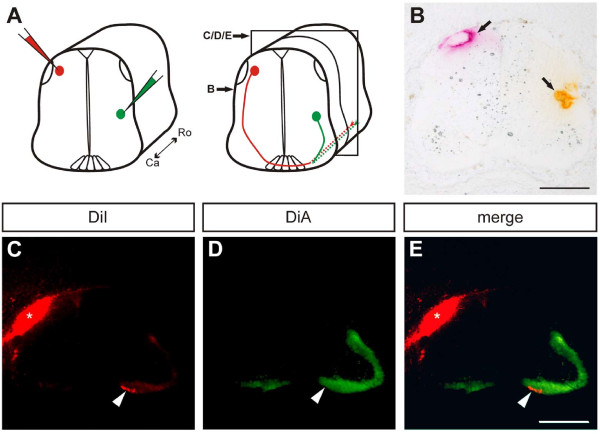
Background: Long-distance axonal growth relies on the precise interplay of guidance cues and cell adhesion molecules. While guidance cues provide positional and directional information for the advancing growth cone, cell adhesion molecules are essential in enabling axonal advancement. Such a dependence on adhesion as well as guidance molecules can be well observed in dorsal commissural interneurons, which follow a highly stereotypical growth and guidance pattern. The mechanisms and molecules involved in the attraction and outgrowth towards the ventral midline, the axon crossing towards the contralateral side, the rostral turning after midline crossing as well as the guidance along the longitudinal axis have been intensely studied. However, little is known about molecules that provide the basis for commissural axon growth along the anterior-posterior axis.
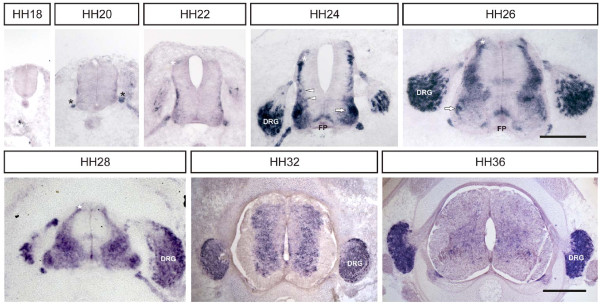
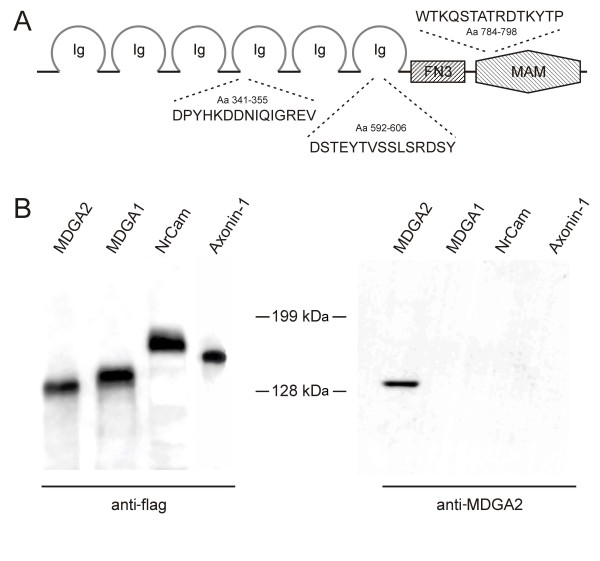
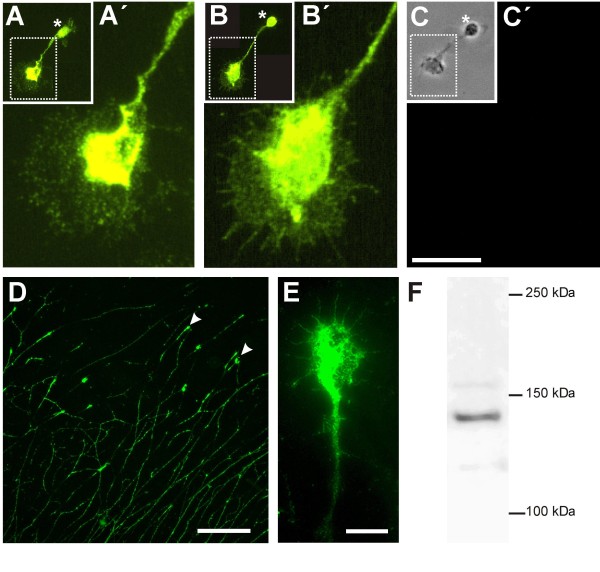
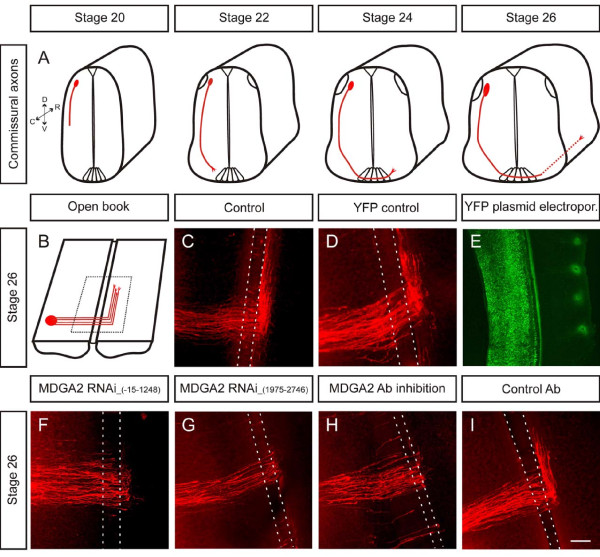
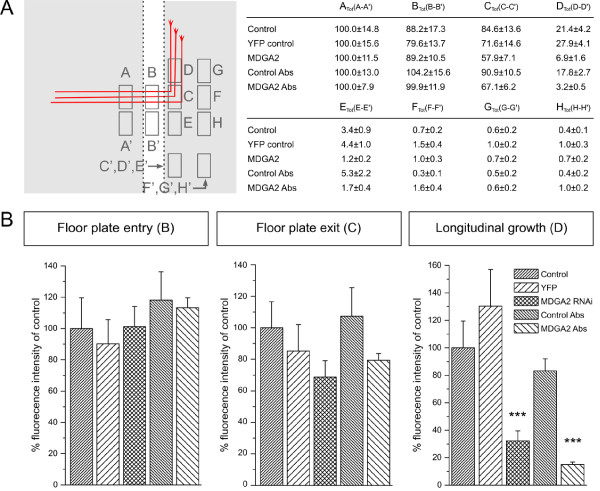
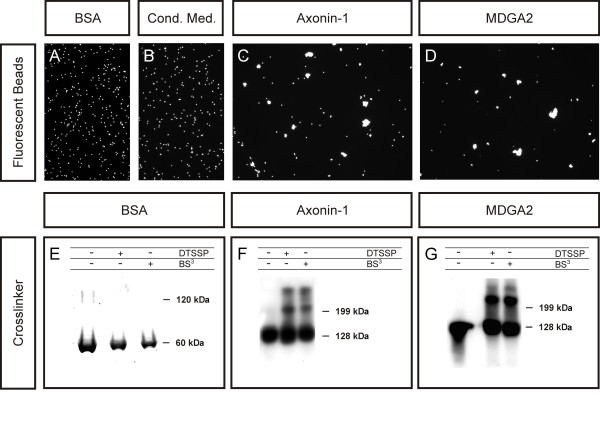
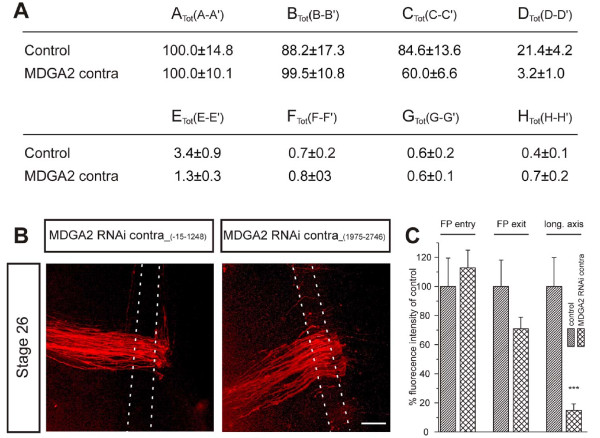
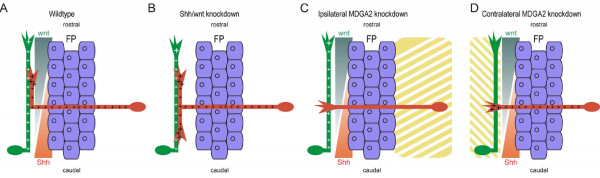
Results: MDGA2, a recently discovered cell adhesion molecule of the IgCAM superfamily, is highly expressed in dorsolaterally located (dI1) spinal interneurons. Functional studies inactivating MDGA2 by RNA interference (RNAi) or function-blocking antibodies demonstrate that either treatment results in a lack of commissural axon growth along the longitudinal axis. Moreover, results from RNAi experiments targeting the contralateral side together with binding studies suggest that homophilic MDGA2 interactions between ipsilaterally projecting axons and post-crossing commissural axons may be the basis of axonal growth along the longitudinal axis.
Conclusions: Directed axonal growth of dorsal commissural interneurons requires an elaborate mixture of instructive (guidance) and permissive (outgrowth supporting) molecules. While Wnt and Sonic hedgehog (Shh) signalling pathways have been shown to specify the growth direction of post-crossing commissural axons, our study now provides evidence that homophilic MDGA2 interactions are essential for axonal extension along the longitudinal axis. Interestingly, so far each part of the complex axonal trajectory of commissural axons uses its own set of guidance and growth-promoting molecules, possibly explaining why such a high number of molecules influencing the growth pattern of commissural interneurons has been identified.
Figures
Similar articles
-
Sonic hedgehog guides commissural axons along the longitudinal axis of the spinal cord.Nat Neurosci. 2005 Mar;8(3):297-304. doi: 10.1038/nn1396. Epub 2005 Jan 30. Nat Neurosci. 2005. PMID: 15746914
-
The Nogo-66 Receptors NgR1 and NgR3 Are Required for Commissural Axon Pathfinding.J Neurosci. 2022 May 18;42(20):4087-4100. doi: 10.1523/JNEUROSCI.1390-21.2022. Epub 2022 Apr 18. J Neurosci. 2022. PMID: 35437280 Free PMC article.
-
Mis-expression of L1 on pre-crossing spinal commissural axons disrupts pathfinding at the ventral midline.Mol Cell Neurosci. 2007 Dec;36(4):462-71. doi: 10.1016/j.mcn.2007.08.003. Epub 2007 Aug 15. Mol Cell Neurosci. 2007. PMID: 17884558 Free PMC article.
-
The spinal cord shows the way - How axons navigate intermediate targets.Dev Biol. 2017 Dec 1;432(1):43-52. doi: 10.1016/j.ydbio.2016.12.002. Epub 2016 Dec 10. Dev Biol. 2017. PMID: 27965053 Review.
-
Axon guidance at the midline choice point.Dev Dyn. 2001 Jun;221(2):154-81. doi: 10.1002/dvdy.1143. Dev Dyn. 2001. PMID: 11376484 Review.
Cited by
-
A Pooling Genome-Wide Association Study Combining a Pathway Analysis for Typical Sporadic Parkinson's Disease in the Han Population of Chinese Mainland.Mol Neurobiol. 2016 Sep;53(7):4302-18. doi: 10.1007/s12035-015-9331-y. Epub 2015 Jul 31. Mol Neurobiol. 2016. PMID: 26227905
-
Mdga2 deficiency leads to an aberrant activation of BDNF/TrkB signaling that underlies autism-relevant synaptic and behavioral changes in mice.PLoS Biol. 2025 Apr 1;23(4):e3003047. doi: 10.1371/journal.pbio.3003047. eCollection 2025 Apr. PLoS Biol. 2025. PMID: 40168357 Free PMC article.
-
Sustained axon regeneration induced by co-deletion of PTEN and SOCS3.Nature. 2011 Nov 6;480(7377):372-5. doi: 10.1038/nature10594. Nature. 2011. PMID: 22056987 Free PMC article.
-
Proper migration and axon outgrowth of zebrafish cranial motoneuron subpopulations require the cell adhesion molecule MDGA2A.Biol Open. 2015 Jan 8;4(2):146-54. doi: 10.1242/bio.20148482. Biol Open. 2015. PMID: 25572423 Free PMC article.
-
Canonical wnt signaling is required for commissural axon guidance.Dev Neurobiol. 2016 Feb;76(2):190-208. doi: 10.1002/dneu.22307. Epub 2015 Jul 3. Dev Neurobiol. 2016. PMID: 26014644 Free PMC article.
References
-
- Stoeckli ET. Molecular mechanisms of commissural axon pathfinding. Prog Brain Res. 1998;117:105–114. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
