

The budding yeast Mei5-Sae3 complex interacts with Rad51 and preferentially binds a DNA fork structure
- PMID: 21543267
- PMCID: PMC3119066
- DOI: 10.1016/j.dnarep.2011.03.006
The budding yeast Mei5-Sae3 complex interacts with Rad51 and preferentially binds a DNA fork structure
Abstract
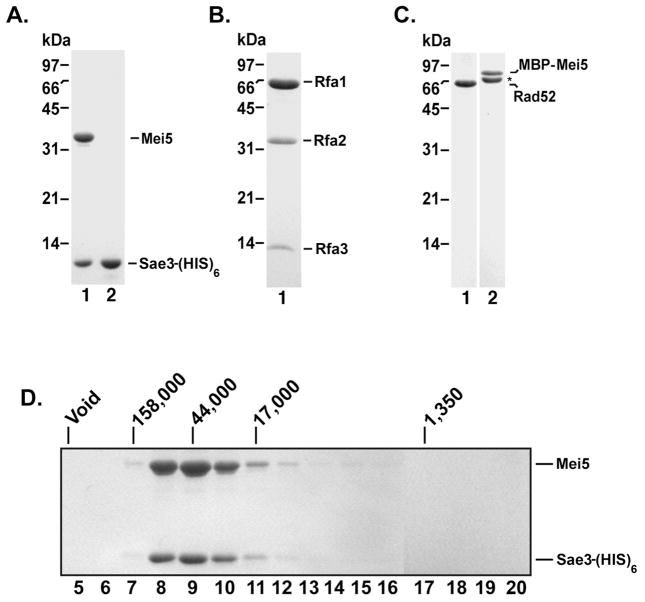
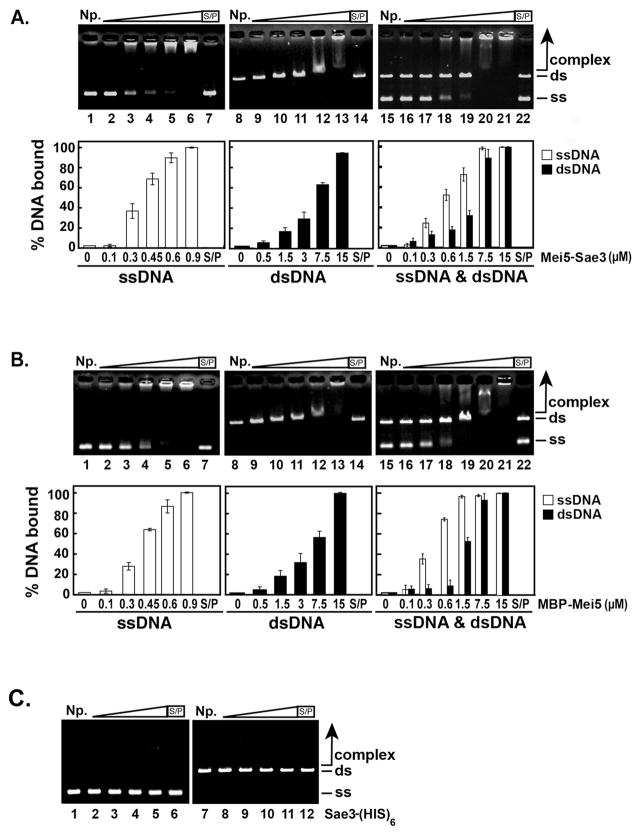
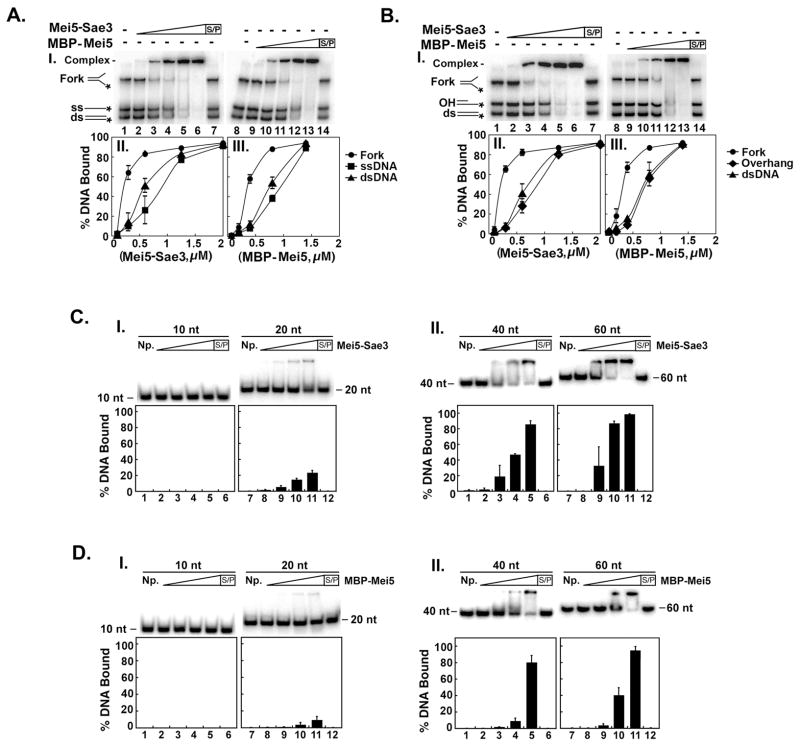
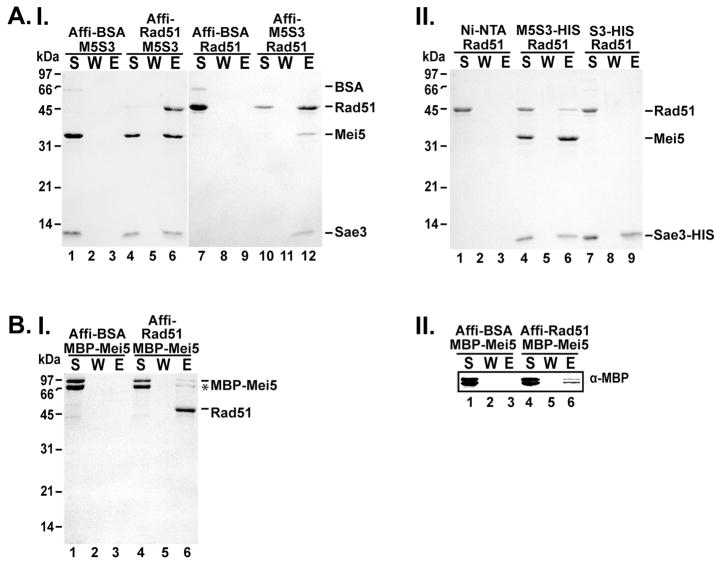
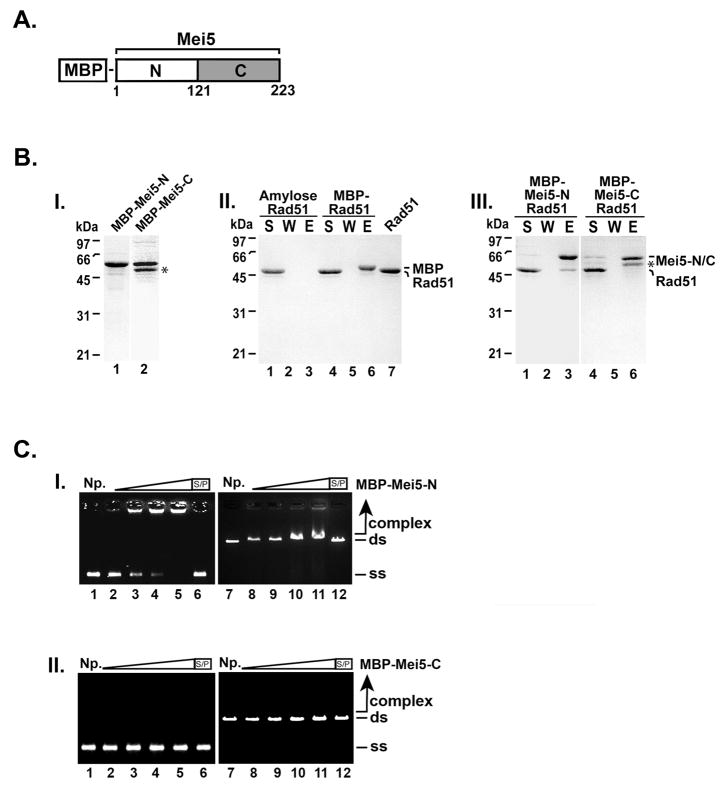
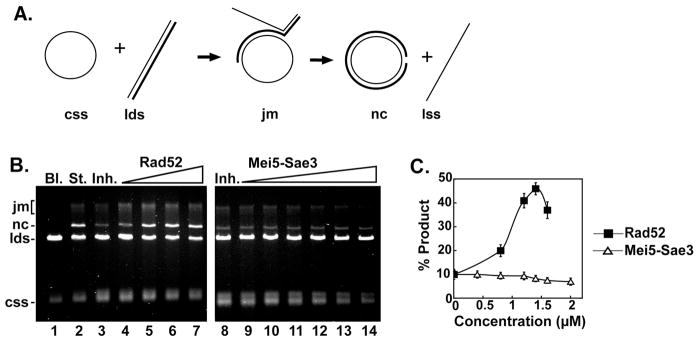
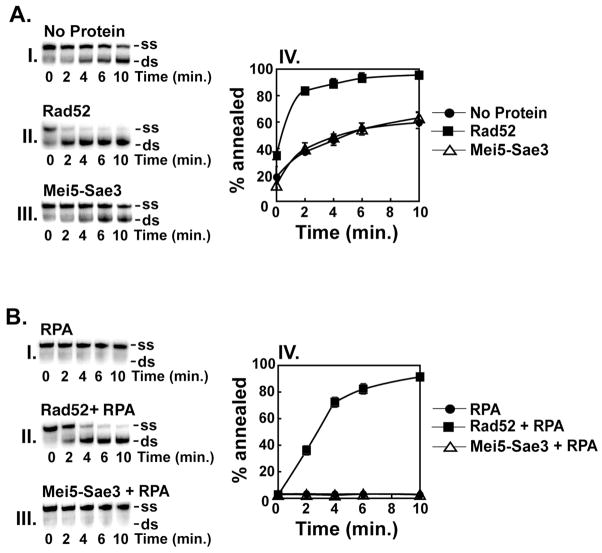
Meiotic homologous recombination in Saccharomyces cerevisiae involves formation of nucleoprotein filaments of Rad51 and Dmc1 that mediate DNA strand exchange between homologous chromosomes. The Mei5-Sae3 protein complex functions as a recombination mediator to promote nucleation of the Dmc1 recombinase onto replication protein A-coated single-stranded DNA. Here, we have expressed and purified the Mei5 protein, Sae3 protein and the Mei5-Sae3 complex for biochemical studies. We show the Mei5-Sae3 complex preferentially binds a fork-like DNA substrate to 3' overhanging DNA, single-stranded DNA or double-stranded DNA. We demonstrate that Mei5 confers DNA binding activity to the Mei5-Sae3 complex. We determined Mei5-Sae3 interacts with the Rad51 recombinase through the N-terminal domain of Mei5. Unlike Rad52, Mei5-Sae3 lacks recombination mediator activity for Rad51. Importantly, we find that the Mei5-Sae3 complex does not harbor single-strand DNA annealing activity. These properties of the Mei5-Sae3 complex distinguishes it from the Rad52 protein, which serves as the mediator of Rad51 and is involved in the single-strand DNA annealing pathway of homologous recombination.
Copyright © 2011 Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
References
-
- Keeney S, Giroux CN, Kleckner N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell. 1997;88:375–384. - PubMed
-
- Shinohara A, Ogawa H, Ogawa T. Rad51 protein involved in repair and recombination in S. cerevisiae is a RecA-like protein. Cell. 1992;69:457–470. - PubMed
-
- Bishop DK, Park D, Xu L, Kleckner N. DMC1: a meiosis-specific yeast homolog of E. coli recA required for recombination, synaptonemal complex formation, and cell cycle progression. Cell. 1992;69:439–456. - PubMed
-
- San Filippo J, Sung P, Klein H. Mechanism of eukaryotic homologous recombination. Annu Rev Biochem. 2008;77:229–257. - PubMed
-
- Sung P, Klein H. Mechanism of homologous recombination: mediators and helicases take on regulatory functions. Nat Rev Mol Cell Biol. 2006;7:739–750. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
