

Identification of amino acid residues important for heparan sulfate proteoglycan interaction within variable region 3 of the feline immunodeficiency virus surface glycoprotein
- PMID: 21543468
- PMCID: PMC3126603
- DOI: 10.1128/JVI.00573-11
Identification of amino acid residues important for heparan sulfate proteoglycan interaction within variable region 3 of the feline immunodeficiency virus surface glycoprotein
Abstract
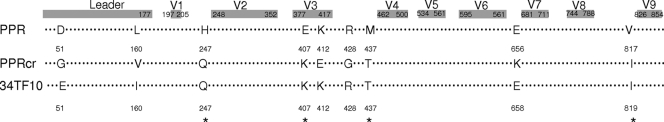
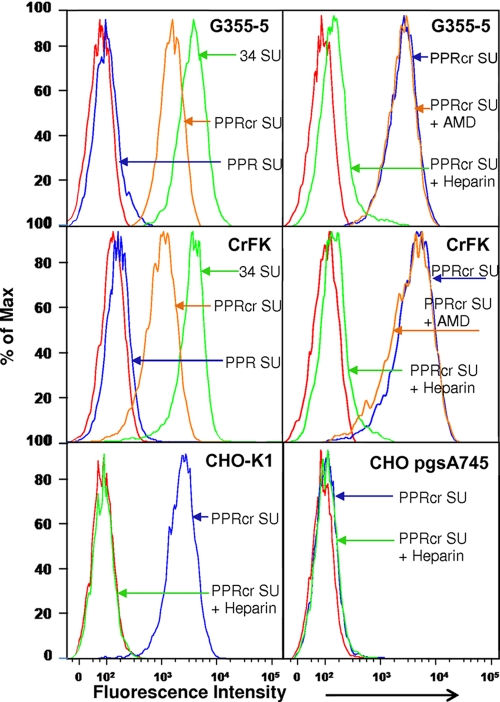
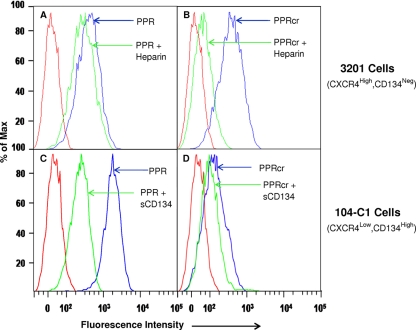
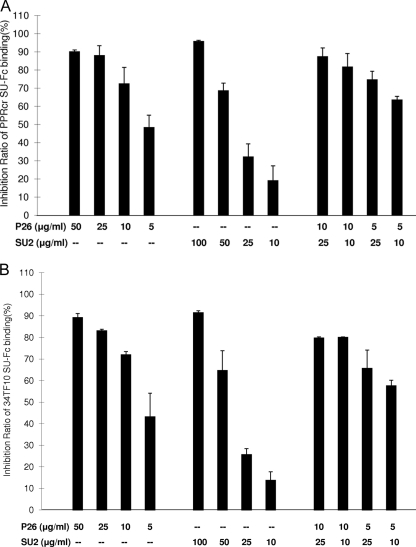
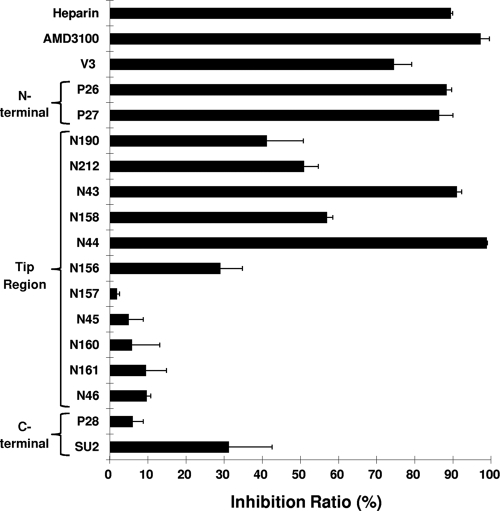
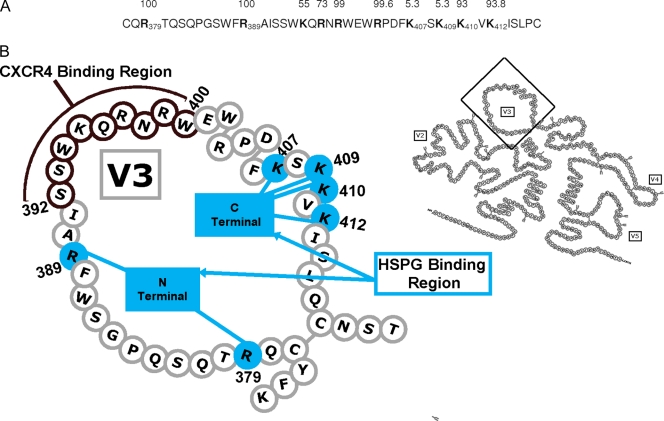
Heparan sulfate proteoglycans (HSPGs) act as binding receptors or attachment factors for the viral envelope of many viruses, including strains of HIV and feline immunodeficiency virus (FIV). The FIV gp95 glycoprotein (SU) from laboratory-adapted strains (tissue culture adapted [TCA]) such as FIV-34TF10 can bind to HSPG, whereas SU from field strains (FS) such as FIV-PPR cannot. Previous studies indicate that SU-HSPG interactions occur within the V3 loop. We utilized a series of nested V3 peptides to further map the HSPG binding sites and found that both sides of the predicted V3 loop stem were critical for the binding but not the CXCR4 binding domain near the predicted tip of the V3 loop. Neutralization assays for TCA strain entry using the same set of V3 peptides showed that peptides targeting CXCR4 or HSPG binding sites can block infection, supporting the V3 loop as a critical neutralization target. Site-directed mutagenesis identified two highly conserved arginines, R379 and R389, on the N-terminal side of the V3 stem as critical for the contact between SU and HSPG. Residues K407, K409, K410, and K412 on the C-terminal side of the V3 stem form a second nonconserved domain necessary for HSPG binding, consistent with the observed specificity distinctions with FS FIV. Our findings discriminate structural determinants important for HSPG and CXCR4 binding by FIV SU and thus further define the importance of the V3 loop for virus entry and infection.
Figures
References
-
- Andrianov A. M., Veresov V. G. 2007. Structural analysis of the HIV-1 gp120 V3 loop: application to the HIV-Haiti isolates. J. Biomol. Struct. Dyn. 24:597–608 - PubMed
-
- Bobardt M. D., et al. 2003. Syndecan captures, protects, and transmits HIV to T lymphocytes. Immunity 18:27–39 - PubMed
-
- Cardin A. D., Weintraub H. J. 1989. Molecular modeling of protein-glycosaminoglycan interactions. Arteriosclerosis 9:21–32 - PubMed
-
- de Parseval A., et al. 2005. A highly conserved arginine in gp120 governs HIV-1 binding to both syndecans and CCR5 via sulfated motifs. J. Biol. Chem. 280:39493–39504 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
