

Inhibitor of DNA binding 4 is expressed selectively by single spermatogonia in the male germline and regulates the self-renewal of spermatogonial stem cells in mice
- PMID: 21543770
- PMCID: PMC3142260
- DOI: 10.1095/biolreprod.111.091330
Inhibitor of DNA binding 4 is expressed selectively by single spermatogonia in the male germline and regulates the self-renewal of spermatogonial stem cells in mice
Abstract
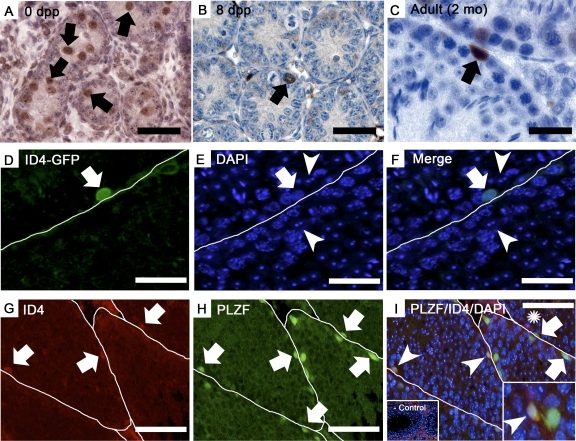
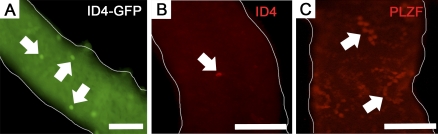
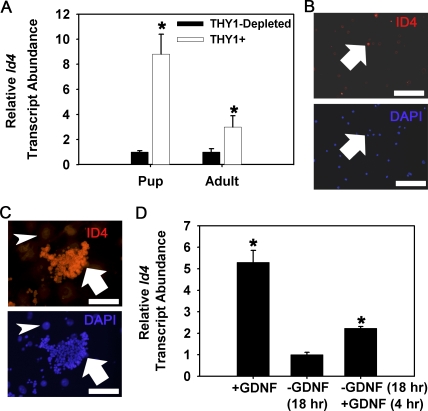
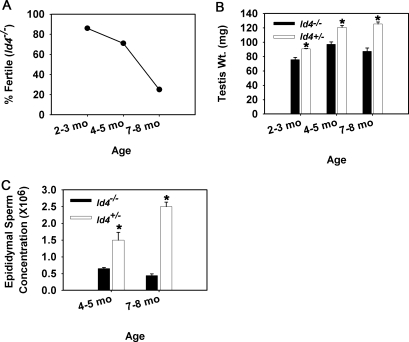
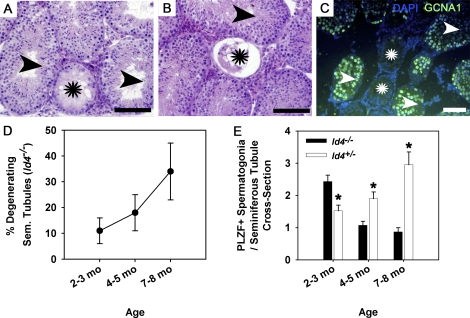
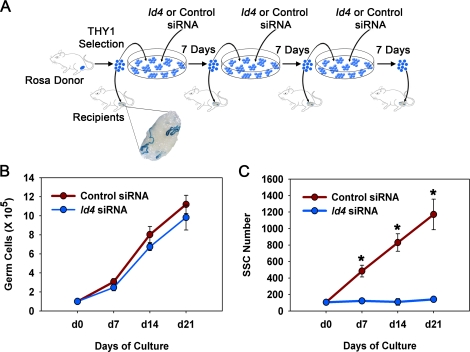
Continual spermatogenesis at a quantitatively normal level is required to sustain male fertility. The foundation of this process relies on maintenance of an undifferentiated spermatogonial population consisting of spermatogonial stem cells (SSCs) that self-renew as well as transient amplifying progenitors produced by differentiation. In mammals, type A(single) spermatogonia form the SSC population, but molecular markers distinguishing these from differentiating progenitors are undefined and knowledge of mechanisms regulating their functions is limited. We show that in the mouse male germline the transcriptional repressor ID4 is expressed by a subpopulation of undifferentiated spermatogonia and selectively marks A(single) spermatogonia. In addition, we found that ID4 expression is up-regulated in isolated SSC-enriched fractions by stimulation from GDNF, a key growth factor driving self-renewal. In mice lacking ID4 expression, quantitatively normal spermatogenesis was found to be impaired due to progressive loss of the undifferentiated spermatogonial population during adulthood. Moreover, reduction of ID4 expression by small interfering RNA treatment abolished the ability of wild-type SSCs to expand in vitro during long-term culture without affecting their survival. Collectively, these results indicate that ID4 is a distinguishing marker of SSCs in the mammalian germline and plays an important role in the regulation of self-renewal.
Figures
Comment in
-
The elusive spermatogonial stem cell marker?Biol Reprod. 2011 Aug;85(2):221-3. doi: 10.1095/biolreprod.111.093146. Epub 2011 May 12. Biol Reprod. 2011. PMID: 21565995 Free PMC article.
References
-
- de Rooij DG, Russell LD. All you wanted to know about spermatogonia but were afraid to ask. J Androl 2000; 21: 776 798. - PubMed
-
- Huckins C. The spermatogonial stem cell population in adult rats. I. Their morphology, proliferation and maturation. Anat Rec 1971; 169: 533 557. - PubMed
-
- Oakberg EF. Spermatogonial stem-cell renewal in the mouse. Anat Rec 1971; 169: 515 531. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
