

Removal of H₂O₂ and generation of superoxide radical: role of cytochrome c and NADH
- PMID: 21545835
- PMCID: PMC3112007
- DOI: 10.1016/j.freeradbiomed.2011.04.007
Removal of H₂O₂ and generation of superoxide radical: role of cytochrome c and NADH
Abstract
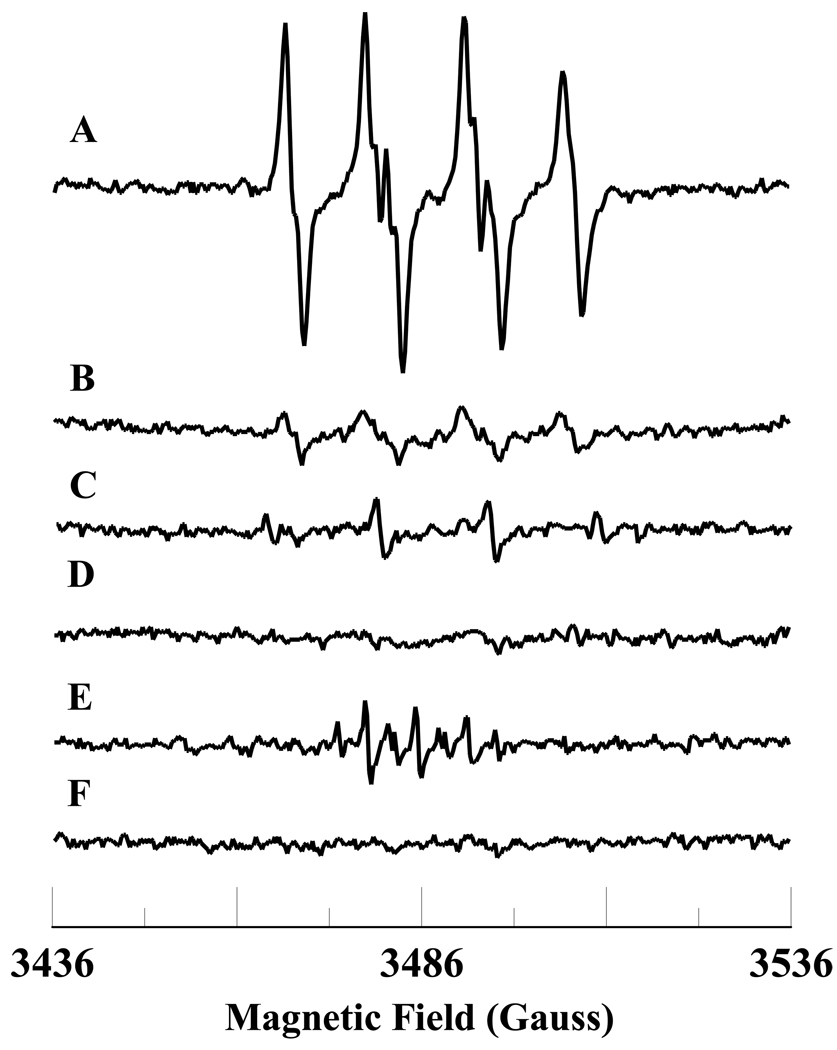
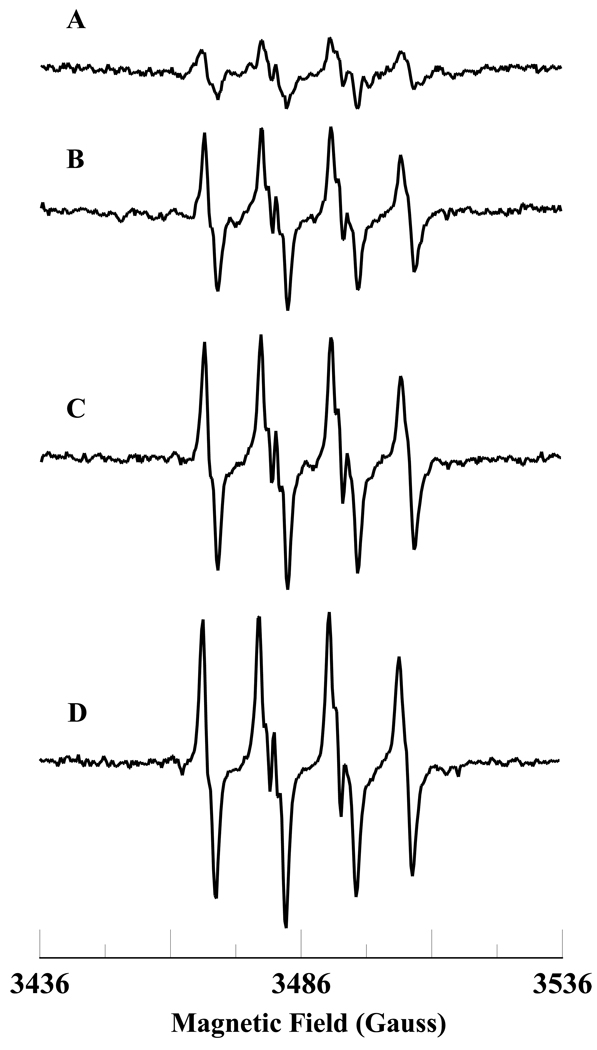
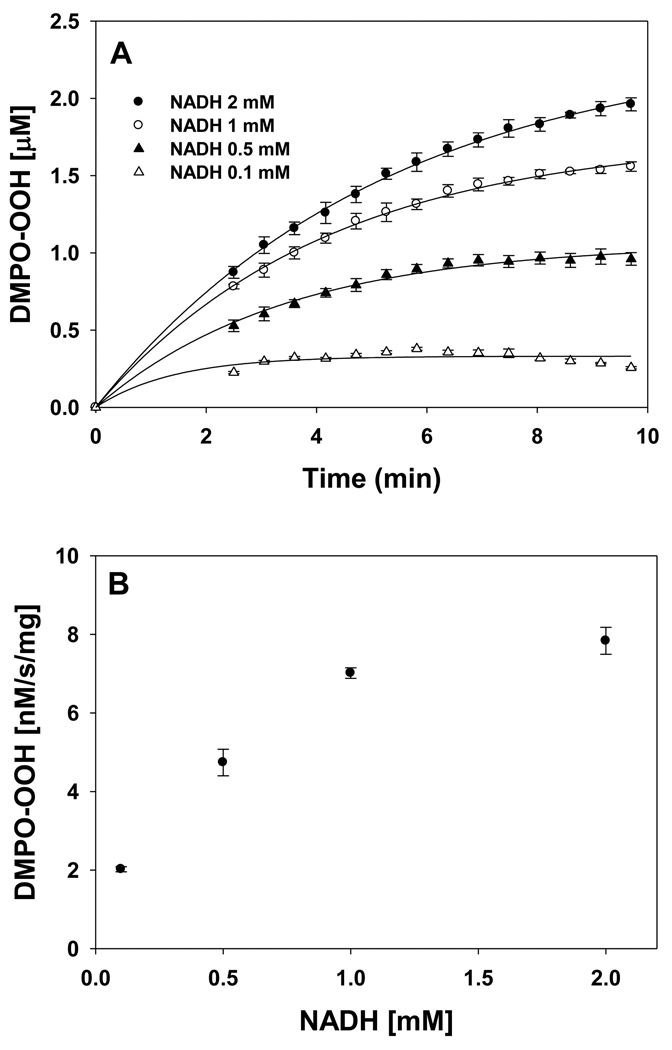
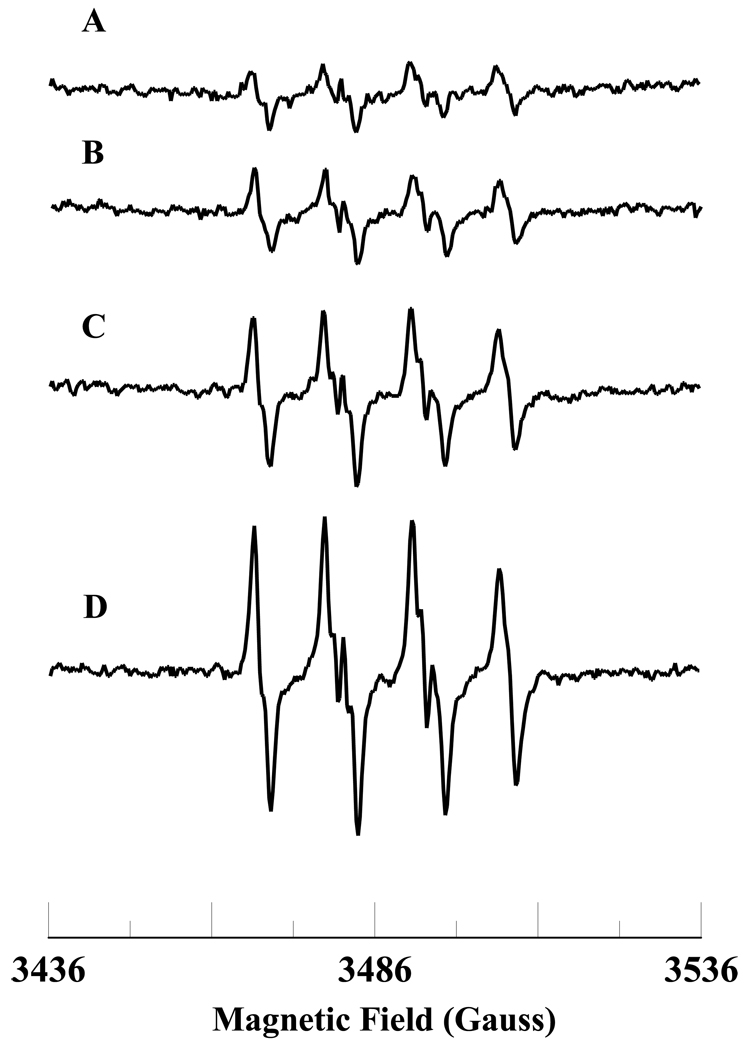
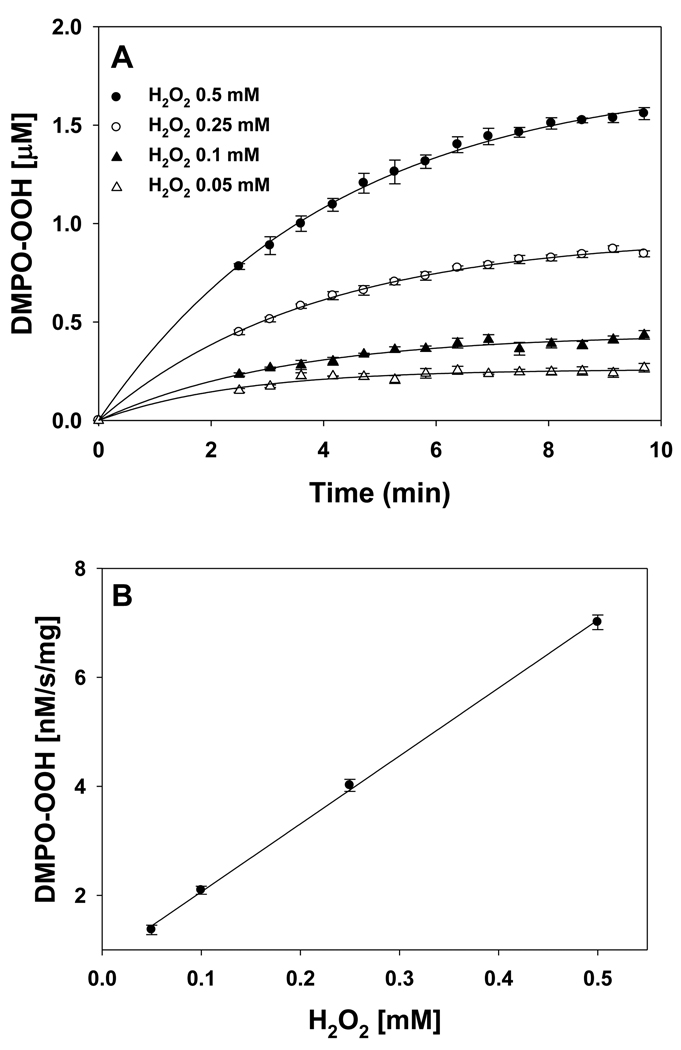
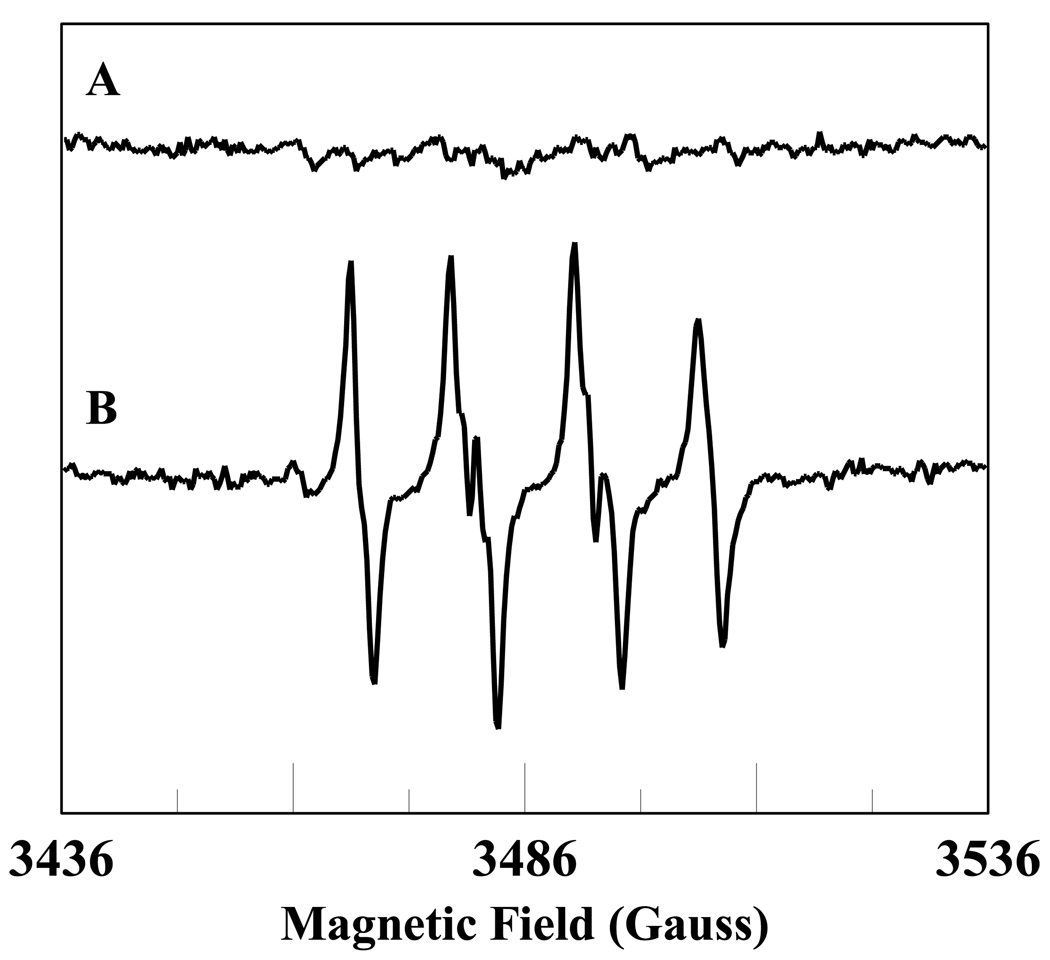
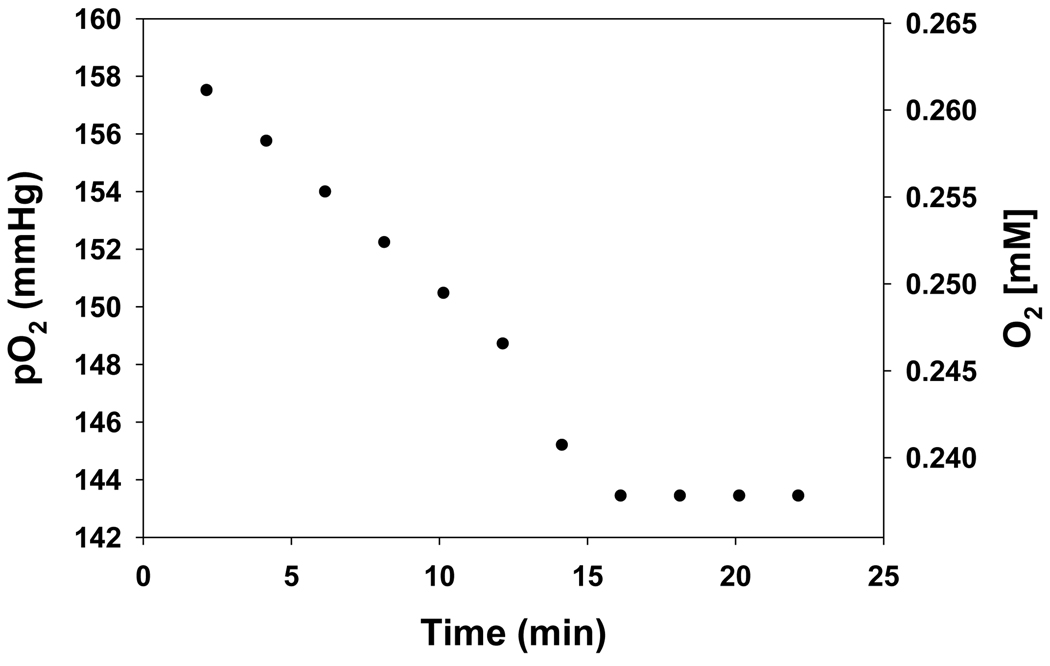
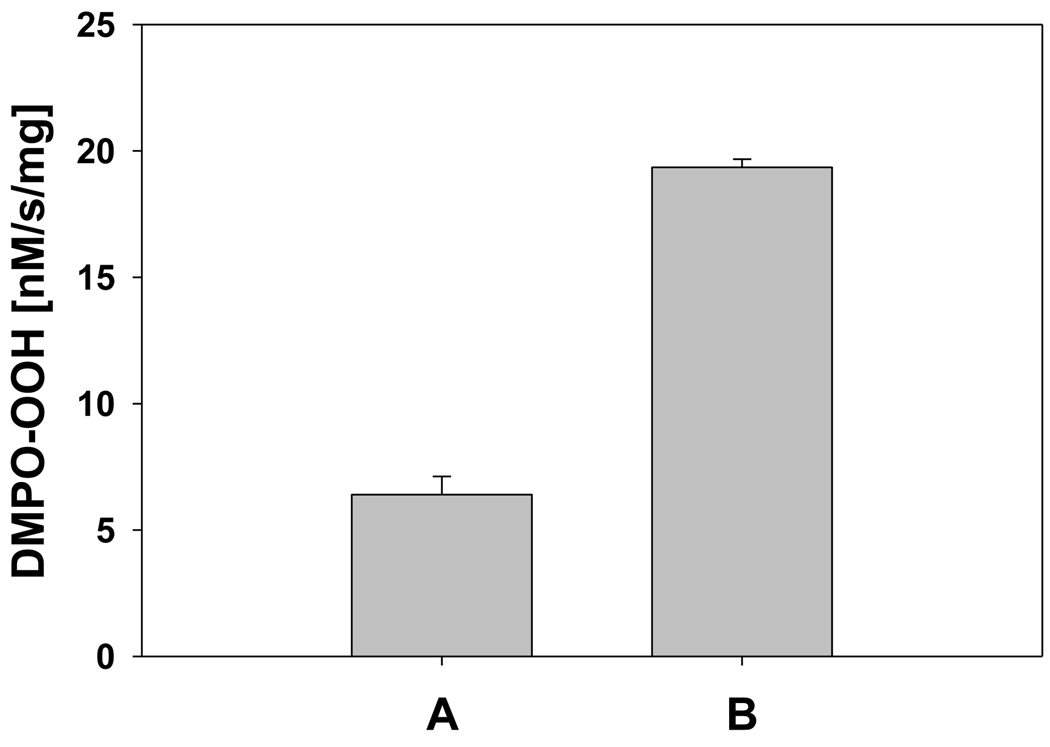
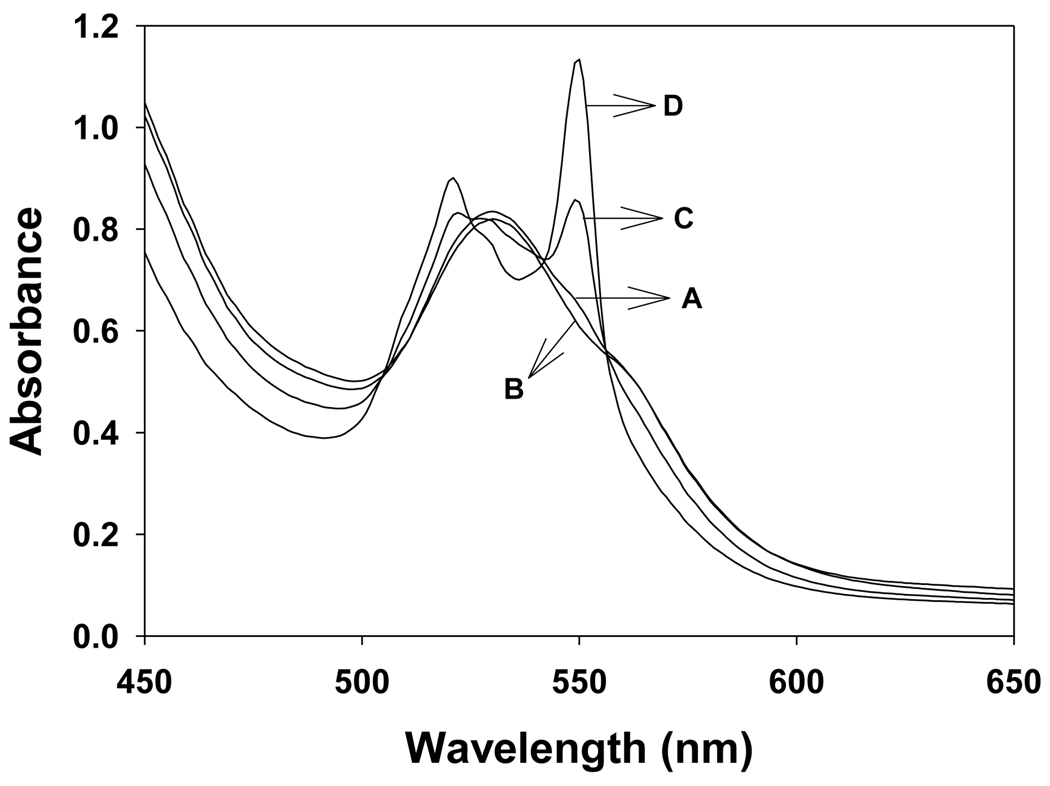
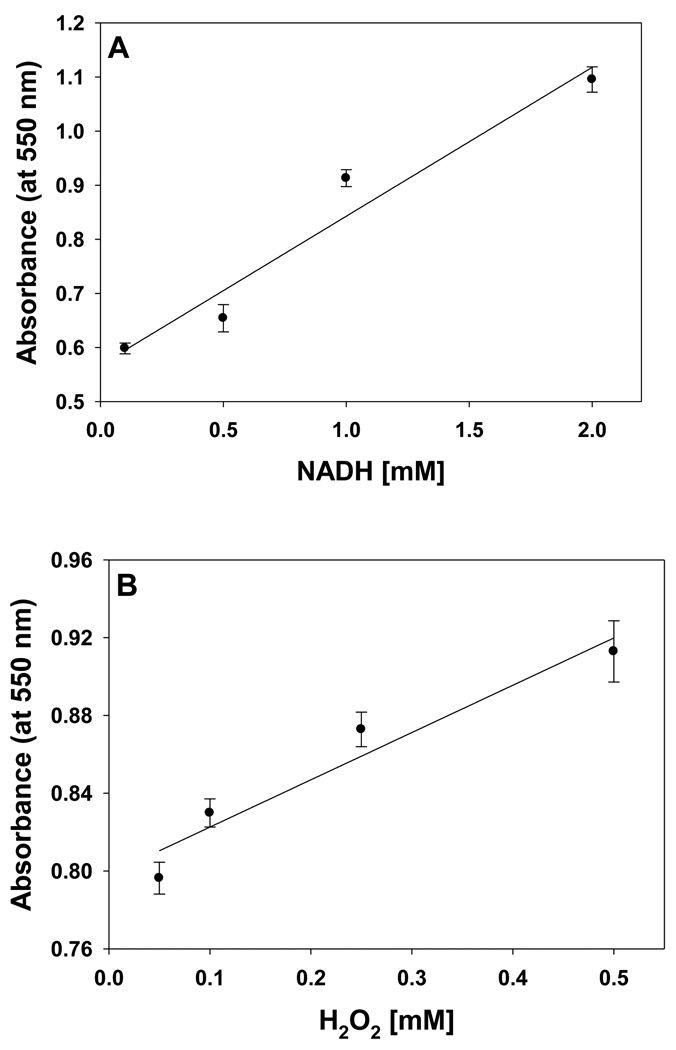
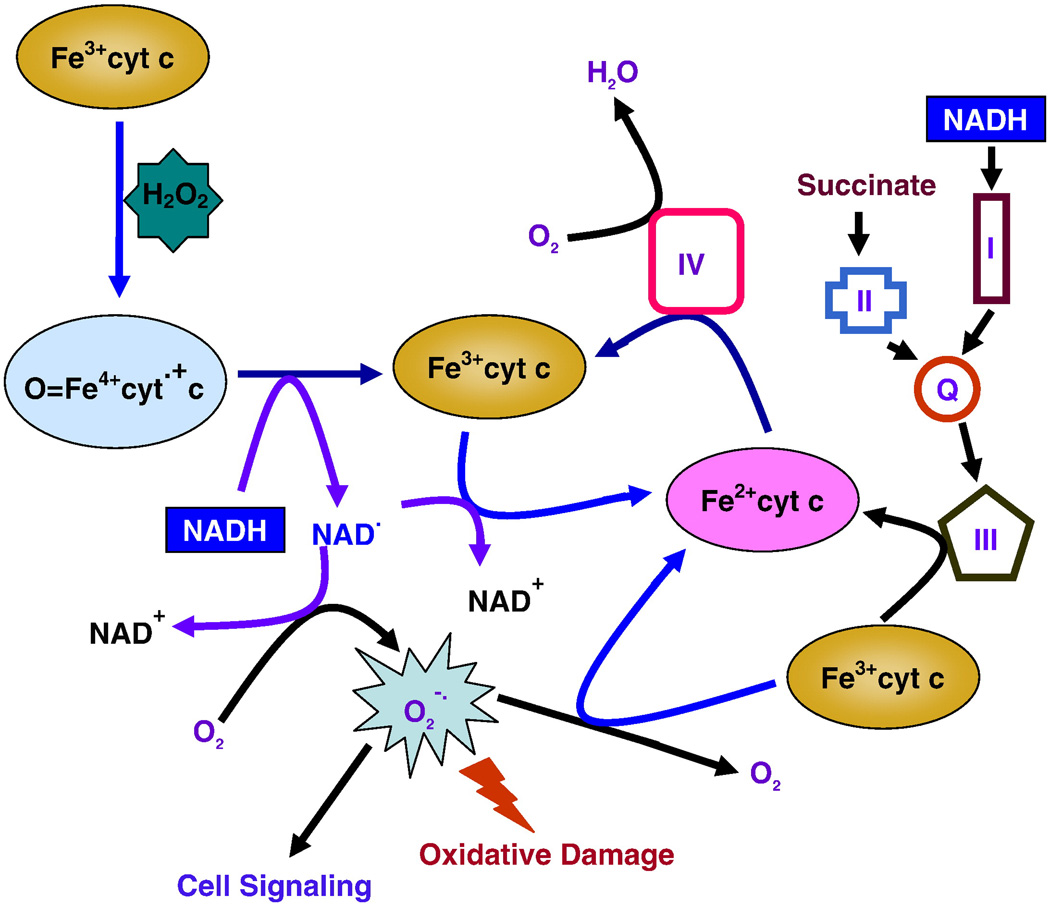
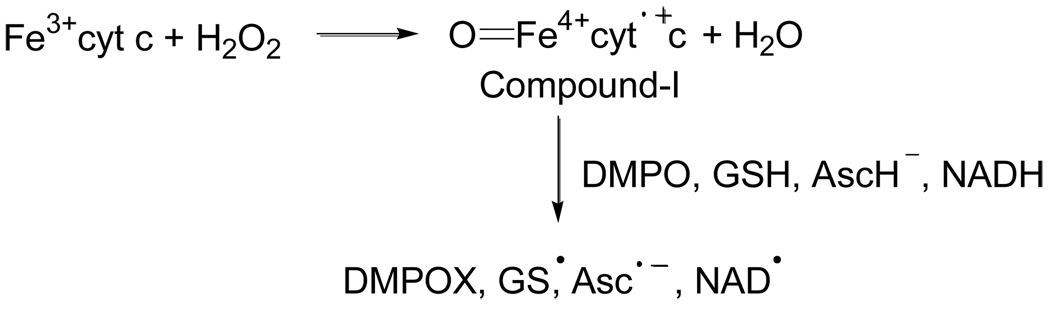
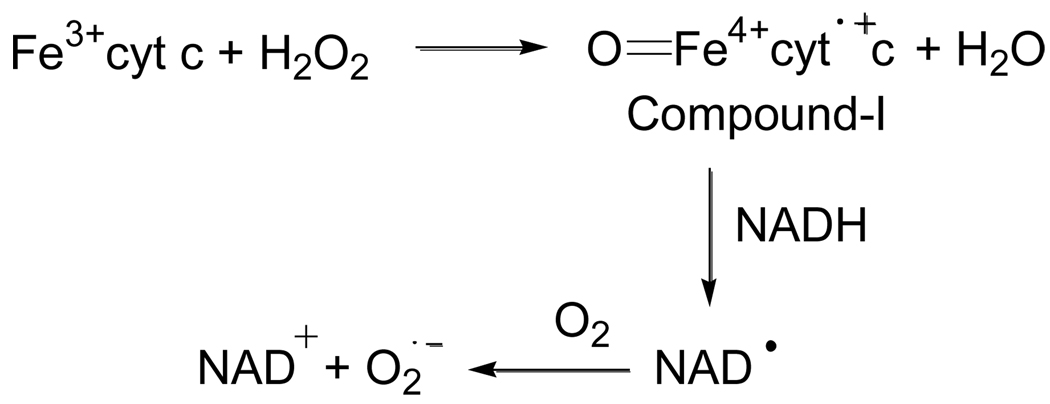
In cells, mitochondria, endoplasmic reticulum, and peroxisomes are the major sources of reactive oxygen species (ROS) under physiological and pathophysiological conditions. Cytochrome c (cyt c) is known to participate in mitochondrial electron transport and has antioxidant and peroxidase activities. Under oxidative or nitrative stress, the peroxidase activity of Fe³⁺cyt c is increased. The level of NADH is also increased under pathophysiological conditions such as ischemia and diabetes and a concurrent increase in hydrogen peroxide (H₂O₂) production occurs. Studies were performed to understand the related mechanisms of radical generation and NADH oxidation by Fe³⁺cyt c in the presence of H₂O₂. Electron paramagnetic resonance (EPR) spin trapping studies using 5,5-dimethyl-1-pyrroline-N-oxide (DMPO) were performed with NADH, Fe³⁺cyt c, and H₂O₂ in the presence of methyl-β-cyclodextrin. An EPR spectrum corresponding to the superoxide radical adduct of DMPO encapsulated in methyl-β-cyclodextrin was obtained. This EPR signal was quenched by the addition of the superoxide scavenging enzyme Cu,Zn-superoxide dismutase (SOD1). The amount of superoxide radical adduct formed from the oxidation of NADH by the peroxidase activity of Fe³⁺cyt c increased with NADH and H₂O₂ concentration. From these results, we propose a mechanism in which the peroxidase activity of Fe³⁺cyt c oxidizes NADH to NAD(•), which in turn donates an electron to O₂, resulting in superoxide radical formation. A UV-visible spectroscopic study shows that Fe³⁺cyt c is reduced in the presence of both NADH and H₂O₂. Our results suggest that Fe³⁺cyt c could have a novel role in the deleterious effects of ischemia/reperfusion and diabetes due to increased production of superoxide radical. In addition, Fe³⁺cyt c may play a key role in the mitochondrial "ROS-induced ROS-release" signaling and in mitochondrial and cellular injury/death. The increased oxidation of NADH and generation of superoxide radical by this mechanism may have implications for the regulation of apoptotic cell death, endothelial dysfunction, and neurological diseases. We also propose an alternative electron transfer pathway, which may protect mitochondria and mitochondrial proteins from oxidative damage.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- St-Pierre J, Buckingham JA, Roebuck SJ, Brand MD. Topology of superoxide production from different sites in the mitochondrial electron transport chain. J. Biol. Chem. 2002;277:44784–44790. - PubMed
-
- Antonenkov VD, Grunau S, Ohlmeier S, Hiltunen JK. Peroxisomes are oxidative organelles. Antioxid. Redox. Signal. 2010;13:525–537. - PubMed
-
- Baines CP, Kaiser RA, Purcell NH, Blair NS, Osinska H, Hambleton MA, Brunskill EW, Sayen MR, Gottlieb RA, Dorn GW, Robbins J, Molkentin JD. Loss of cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature. 2005;434:658–662. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
