

Trex2 enables spontaneous sister chromatid exchanges without facilitating DNA double-strand break repair
- PMID: 21546543
- PMCID: PMC3176084
- DOI: 10.1534/genetics.111.129833
Trex2 enables spontaneous sister chromatid exchanges without facilitating DNA double-strand break repair
Abstract
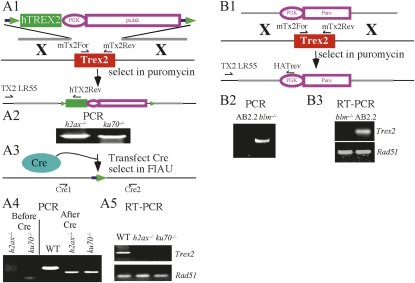
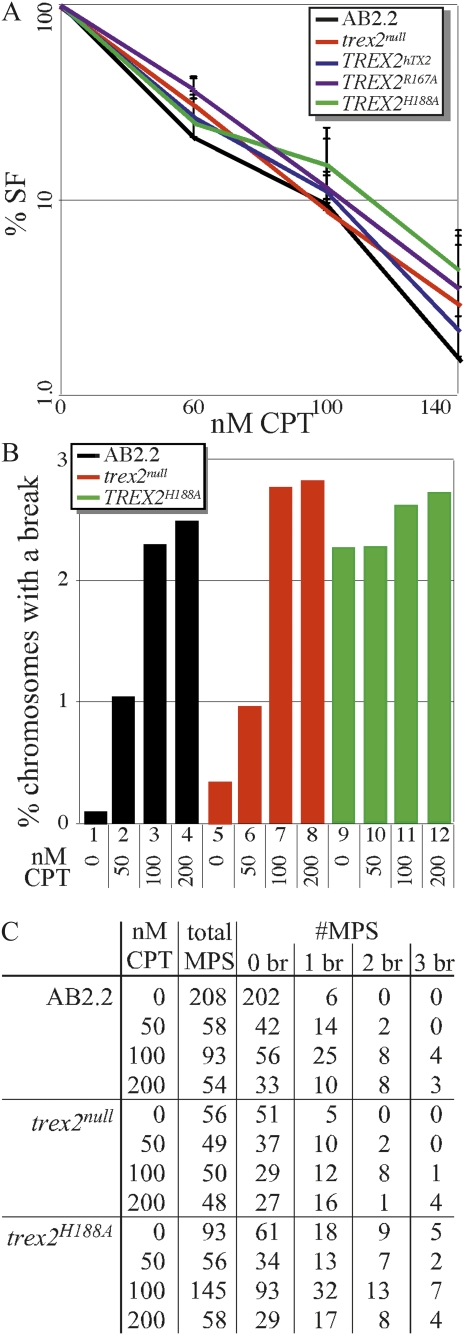
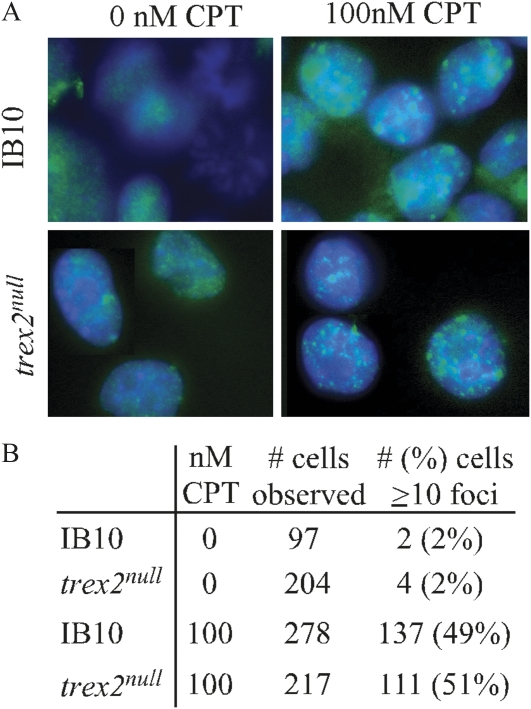
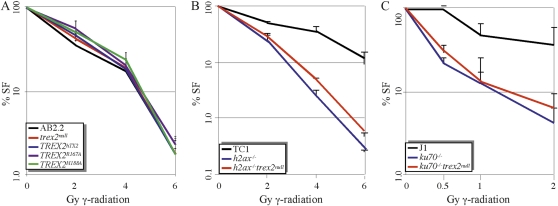
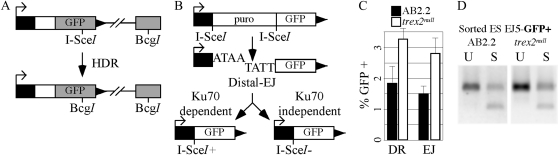
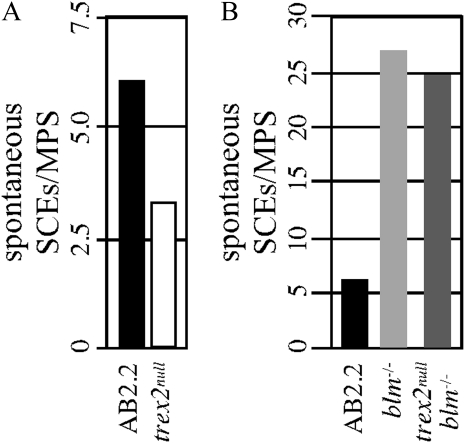
Trex2 is a 3' → 5' exonuclease that removes 3'-mismatched sequences in a biochemical assay; however, its biological function remains unclear. To address biology we previously generated trex2(null) mouse embryonic stem (ES) cells and expressed in these cells wild-type human TREX2 cDNA (Trex2(hTX2)) or cDNA with a single-amino-acid change in the catalytic domain (Trex2(H188A)) or in the DNA-binding domain (Trex2(R167A)). We found the trex2(null) and Trex2(H188A) cells exhibited spontaneous broken chromosomes and trex2(null) cells exhibited spontaneous chromosomal rearrangements. We also found ectopically expressed human TREX2 was active at the 3' ends of I-SceI-induced chromosomal double-strand breaks (DSBs). Therefore, we hypothesized Trex2 participates in DNA DSB repair by modifying 3' ends. This may be especially important for ends with damaged nucleotides. Here we present data that are unexpected and prompt a new model. We found Trex2-altered cells (null, H188A, and R167A) were not hypersensitive to camptothecin, a type-1 topoisomerase inhibitor that induces DSBs at replication forks. In addition, Trex2-altered cells were not hypersensitive to γ-radiation, an agent that causes DSBs throughout the cell cycle. This observation held true even in cells compromised for one of the two major DSB repair pathways: homology-directed repair (HDR) or nonhomologous end joining (NHEJ). Trex2 deletion also enhanced repair of an I-SceI-induced DSB by both HDR and NHEJ without affecting pathway choice. Interestingly, however, trex2(null) cells exhibited reduced spontaneous sister chromatid exchanges (SCEs) but this was not due to a defect in HDR-mediated crossing over. Therefore, reduced spontaneous SCE could be a manifestation of the same defect that caused spontaneous broken chromosomes and spontaneous chromosomal rearrangements. These unexpected data suggest Trex2 does not enable DSB repair and prompt a new model that posits Trex2 suppresses the formation of broken chromosomes.
Figures
Similar articles
-
Two replication fork maintenance pathways fuse inverted repeats to rearrange chromosomes.Nature. 2013 Sep 26;501(7468):569-72. doi: 10.1038/nature12500. Epub 2013 Sep 8. Nature. 2013. PMID: 24013173 Free PMC article.
-
TREX2 exonuclease defective cells exhibit double-strand breaks and chromosomal fragments but not Robertsonian translocations.Mutat Res. 2009 Mar 9;662(1-2):84-7. doi: 10.1016/j.mrfmmm.2008.11.012. Epub 2008 Nov 27. Mutat Res. 2009. PMID: 19094998 Free PMC article.
-
Limiting the persistence of a chromosome break diminishes its mutagenic potential.PLoS Genet. 2009 Oct;5(10):e1000683. doi: 10.1371/journal.pgen.1000683. Epub 2009 Oct 16. PLoS Genet. 2009. PMID: 19834534 Free PMC article.
-
Progress towards understanding the nature of chromatid breakage.Cytogenet Genome Res. 2004;104(1-4):65-71. doi: 10.1159/000077467. Cytogenet Genome Res. 2004. PMID: 15162016 Review.
-
Mechanisms of DNA double strand break repair and chromosome aberration formation.Cytogenet Genome Res. 2004;104(1-4):14-20. doi: 10.1159/000077461. Cytogenet Genome Res. 2004. PMID: 15162010 Review.
Cited by
-
Chromosomal Rearrangements in Cancer: Detection and potential causal mechanisms.Mol Cell Oncol. 2014 Jul;1(1):e29904. doi: 10.4161/mco.29904. Mol Cell Oncol. 2014. PMID: 26203462 Free PMC article.
-
Methods Favoring Homology-Directed Repair Choice in Response to CRISPR/Cas9 Induced-Double Strand Breaks.Int J Mol Sci. 2020 Sep 4;21(18):6461. doi: 10.3390/ijms21186461. Int J Mol Sci. 2020. PMID: 32899704 Free PMC article. Review.
-
Structural insights into the duplex DNA processing of TREX2.Nucleic Acids Res. 2018 Dec 14;46(22):12166-12176. doi: 10.1093/nar/gky970. Nucleic Acids Res. 2018. PMID: 30357414 Free PMC article.
-
Two replication fork maintenance pathways fuse inverted repeats to rearrange chromosomes.Nature. 2013 Sep 26;501(7468):569-72. doi: 10.1038/nature12500. Epub 2013 Sep 8. Nature. 2013. PMID: 24013173 Free PMC article.
-
Defining a genotoxic profile with mouse embryonic stem cells.Exp Biol Med (Maywood). 2013 Mar;238(3):285-93. doi: 10.1177/1535370213480700. Exp Biol Med (Maywood). 2013. PMID: 23598974 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
