

GIV/Girdin is a rheostat that fine-tunes growth factor signals during tumor progression
- PMID: 21546796
- PMCID: PMC3210208
- DOI: 10.4161/cam.5.3.15909
GIV/Girdin is a rheostat that fine-tunes growth factor signals during tumor progression
Abstract
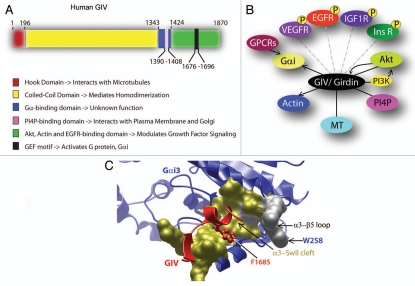
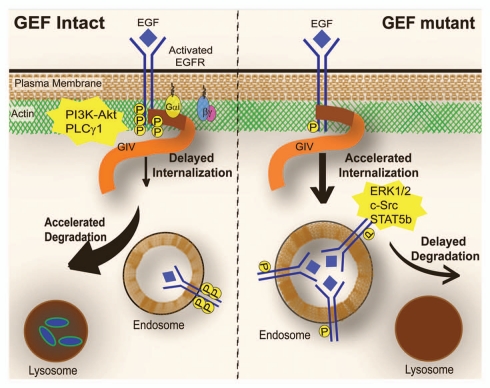
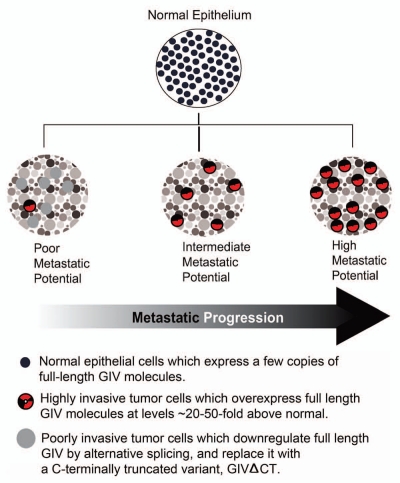
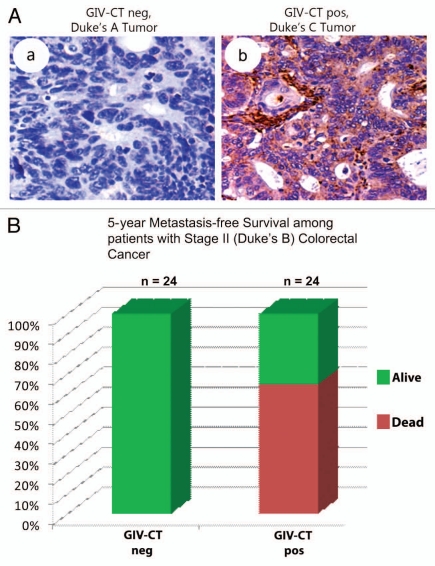
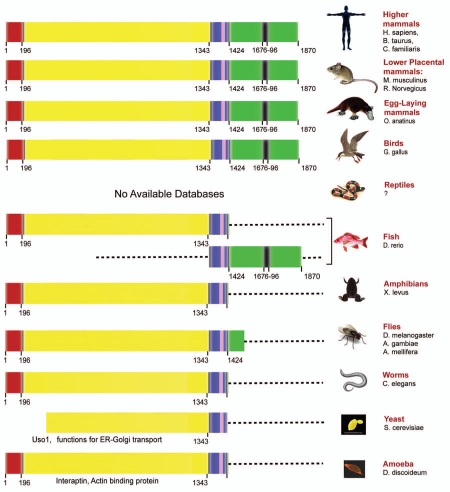
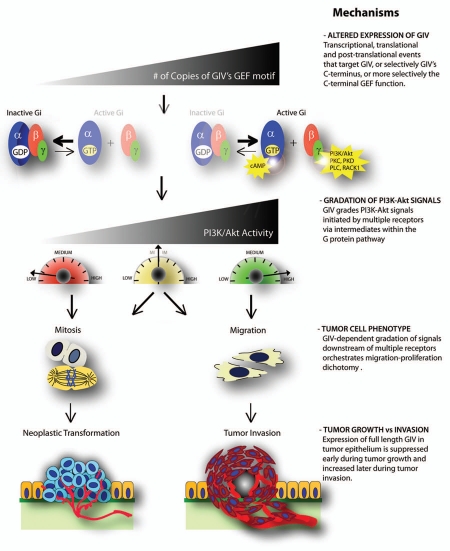
GIV/Girdin is a multidomain signaling molecule that enhances PI3K-Akt signals downstream of both G protein-coupled and growth factor receptors. We previously reported that GIV triggers cell migration via its C-terminal guanine-nucleotide exchange factor (GEF) motif that activates Gαi. Recently we discovered that GIV's C-terminus directly interacts with the epidermal growth factor receptor (EGFR), and when its GEF function is intact, a Gαi-GIV-EGFR signaling complex assembles. By coupling G proteins to growth factor receptors, GIV is uniquely poised to intercept the incoming receptor-initiated signals and modulate them via G protein intermediates. Subsequent work has revealed that expression of the highly specialized C-terminus of GIV undergoes a bipartite dysregulation during oncogenesis-full length GIV with an intact C-terminus is expressed at levels ~20-50-fold above normal in highly invasive cancer cells and metastatic tumors, but its C-terminus is truncated by alternative splicing in poorly invasive cancer cells and non-invasive tumors. The consequences of such dysregulation on graded signal transduction and cellular phenotypes in the normal epithelium and its implication during tumor progression are discussed herein. Based on the fact that GIV grades incoming signals initiated by ligand-activated receptors by linking them to cyclical activation of G proteins, we propose that GIV is a molecular rheostat for signal transduction.
Figures
References
-
- Le-Niculescu H, Niesman I, Fischer T, DeVries L, Farquhar MG. Identification and characterization of GIV, a novel Galpha i/s-interacting protein found on COPI, endoplasmic reticulum-Golgi transport vesicles. J Biol Chem. 2005;280:22012–22020. - PubMed
-
- Simpson F, Martin S, Evans TM, Kerr M, James DE, Parton RG, et al. A novel hook-related protein family and the characterization of hook-related protein 1. Traffic. 2005;6:442–458. - PubMed
-
- Anai M, Shojima N, Katagiri H, Ogihara T, Sakoda H, Onishi Y, et al. A novel protein kinase B (PKB)/AKT-binding protein enhances PKB kinase activity and regulates DNA synthesis. J Biol Chem. 2005;280:18525–18535. - PubMed
-
- Enomoto A, Murakami H, Asai N, Morone N, Watanabe T, Kawai K, et al. Akt/PKB regulates actin organization and cell motility via Girdin/APE. Dev Cell. 2005;9:389–402. - PubMed
-
- Enomoto A, Ping J, Takahashi M. Girdin, a novel actin-binding protein, and its family of proteins possess versatile functions in the Akt and Wnt signaling pathways. Ann NY Acad Sci. 2006;1086:169–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous