

TTBK2 kinase substrate specificity and the impact of spinocerebellar-ataxia-causing mutations on expression, activity, localization and development
- PMID: 21548880
- PMCID: PMC3739326
- DOI: 10.1042/BJ20110276
TTBK2 kinase substrate specificity and the impact of spinocerebellar-ataxia-causing mutations on expression, activity, localization and development
Abstract
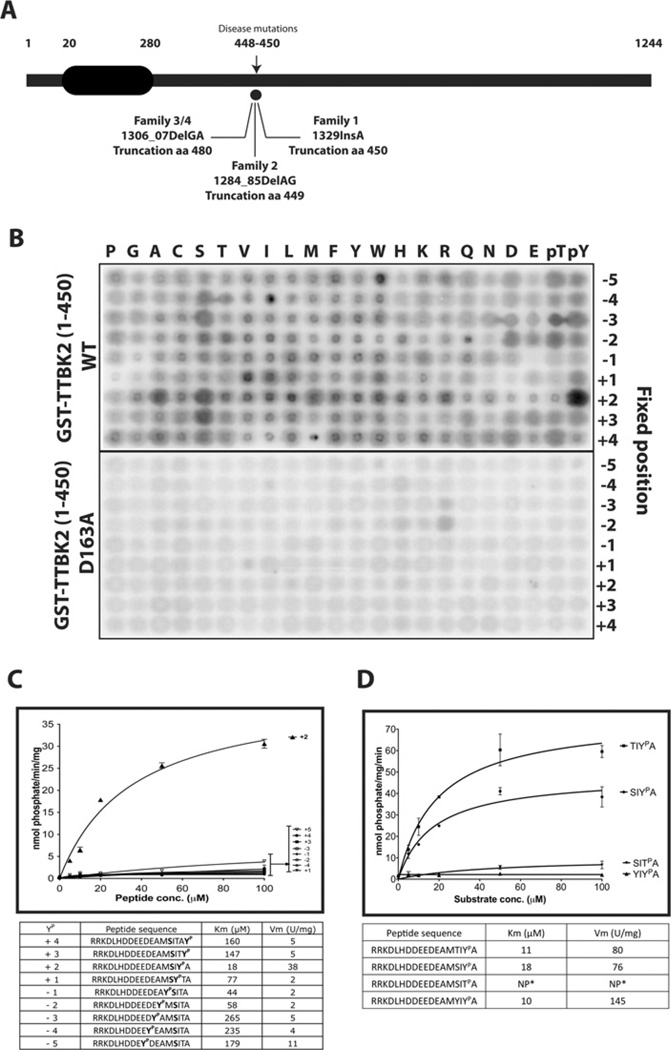
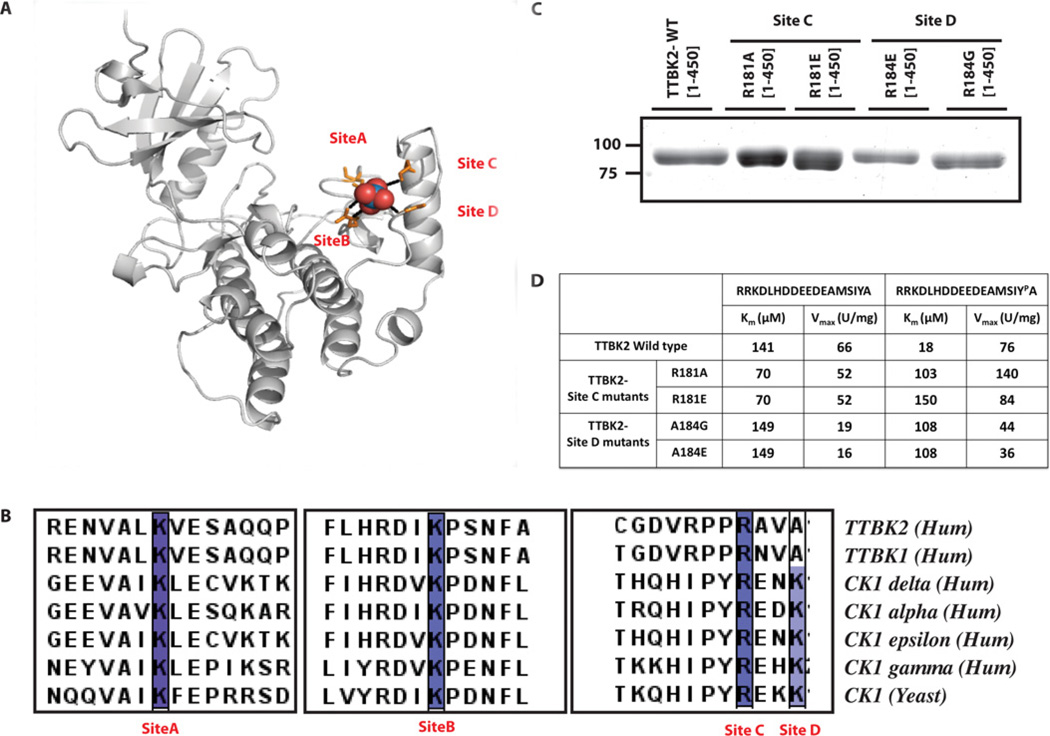
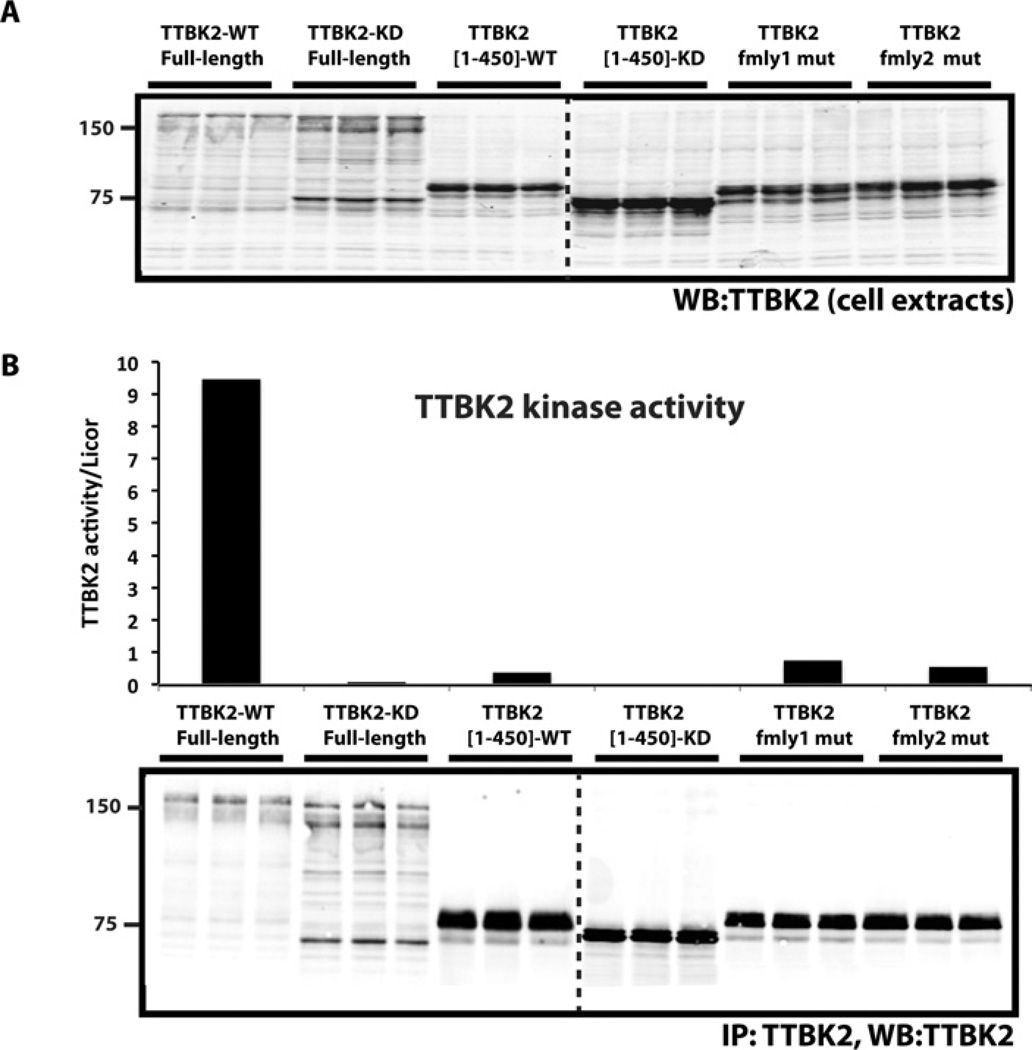
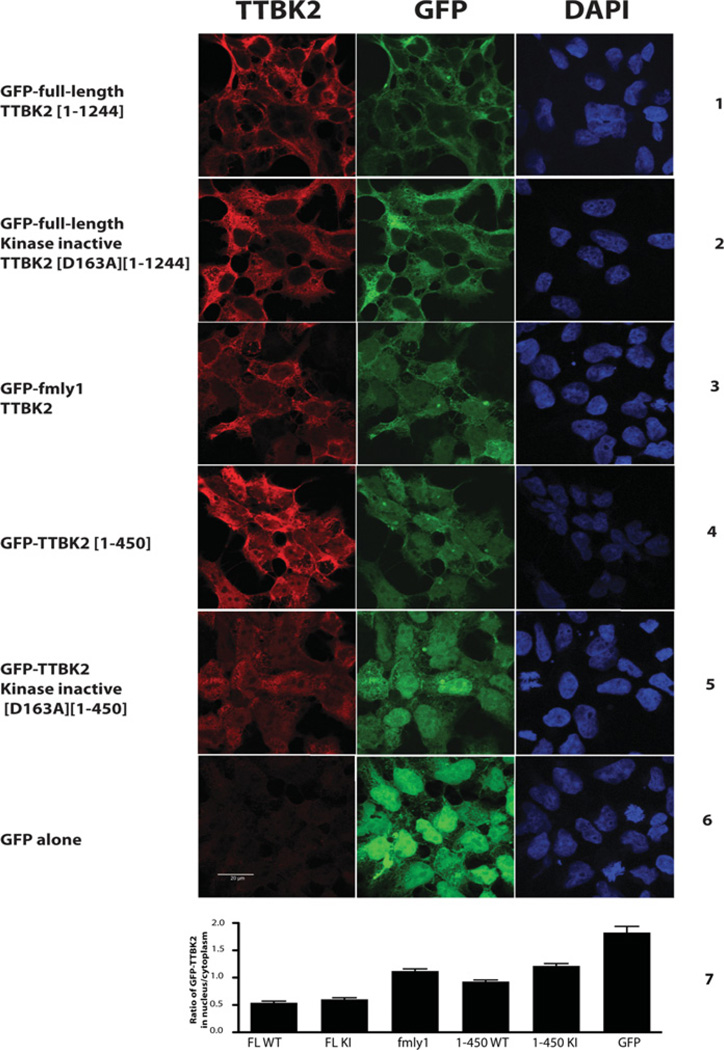
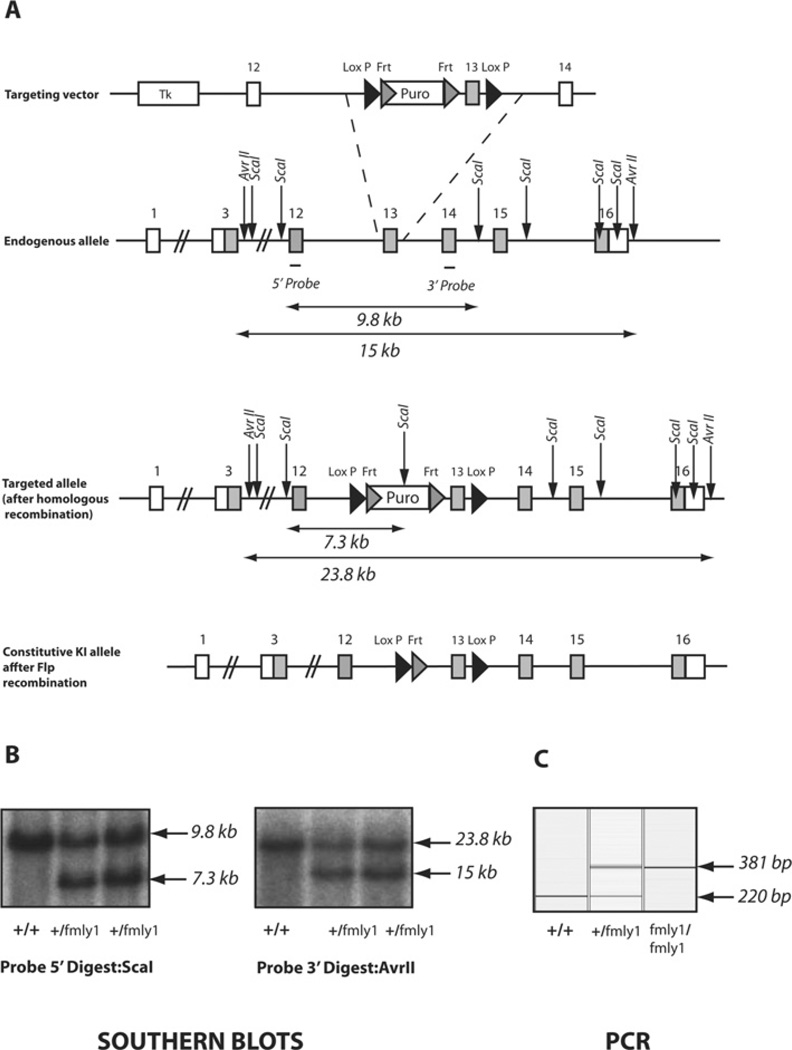
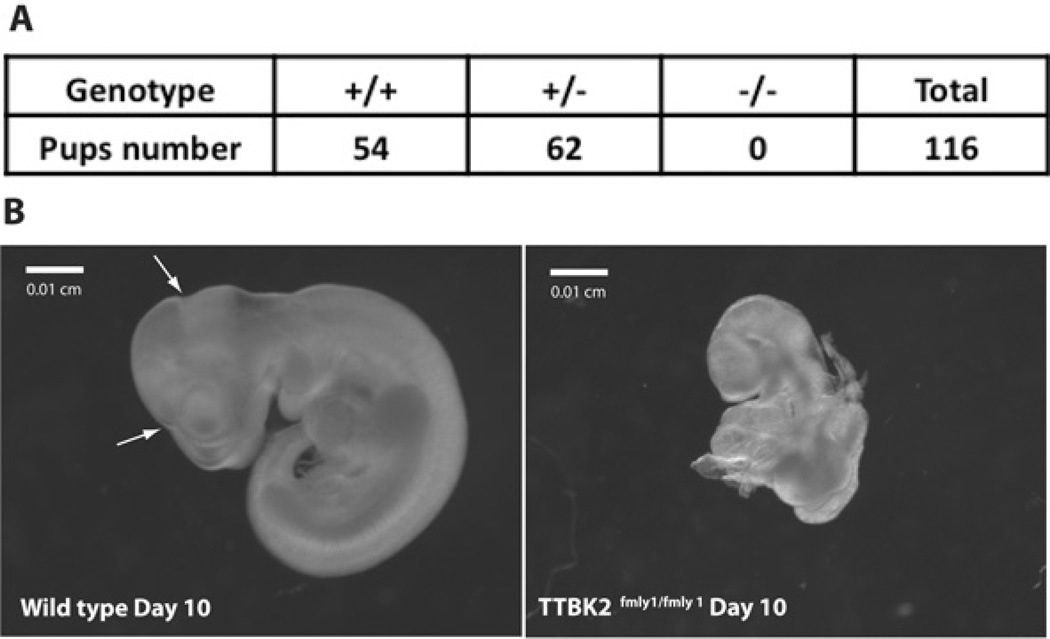
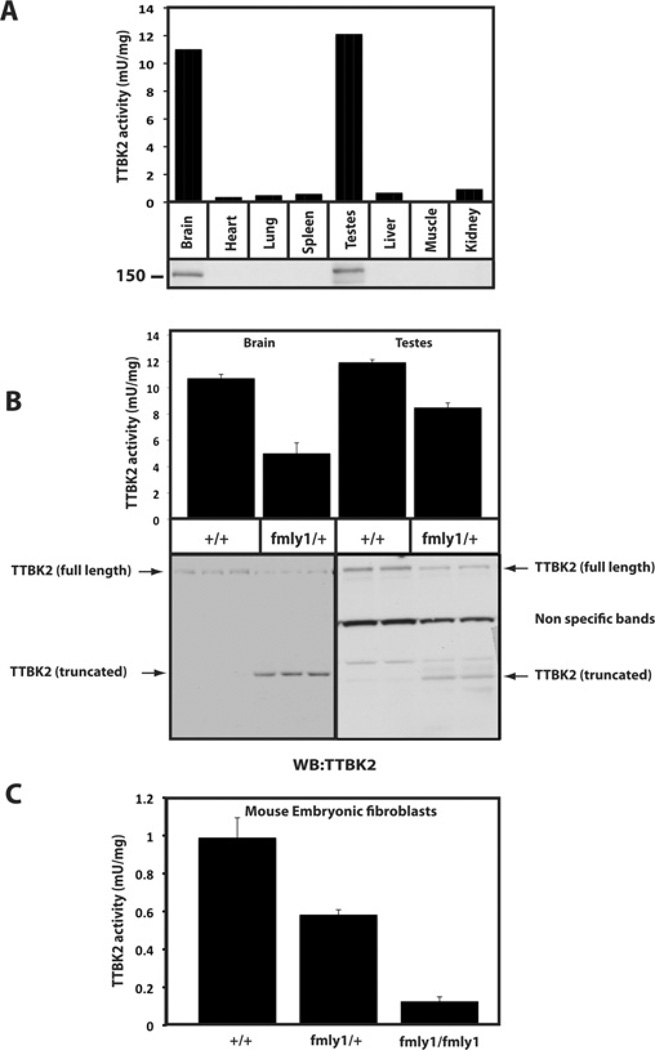
Mutations that truncate the C-terminal non-catalytic moiety of TTBK2 (tau tubulin kinase 2) cause the inherited, autosomal dominant, SCA11 (spinocerebellar ataxia type 11) movement disorder. In the present study we first assess the substrate specificity of TTBK2 and demonstrate that it has an unusual preference for a phosphotyrosine residue at the +2 position relative to the phosphorylation site. We elaborate a peptide substrate (TTBKtide, RRKDLHDDEEDEAMSIYpA) that can be employed to quantify TTBK2 kinase activity. Through modelling and mutagenesis we identify a putative phosphate-priming groove within the TTBK2 kinase domain. We demonstrate that SCA11 truncating mutations promote TTBK2 protein expression, suppress kinase activity and lead to enhanced nuclear localization. We generate an SCA11-mutation-carrying knockin mouse and show that this leads to inhibition of endogenous TTBK2 protein kinase activity. Finally, we find that, in homozygosity, the SCA11 mutation causes embryonic lethality at embryonic day 10. These findings provide the first insights into some of the intrinsic properties of TTBK2 and reveal how SCA11-causing mutations affect protein expression, catalytic activity, localization and development. We hope that these findings will be helpful for future investigation of the regulation and function of TTBK2 and its role in SCA11.
Figures
References
-
- Houlden H, Johnson J, Gardner-Thorpe C, Lashley T, Hernandez D, Worth P, Singleton AB, Hilton DA, Holton J, Revesz T, et al. Mutations in TTBK2, encoding a kinase implicated in tau phosphorylation, segregate with spinocerebellar ataxia type 11. Nat. Genet. 2007;39:1434–1436. - PubMed
-
- Bauer P, Stevanin G, Beetz C, Synofzik M, Schmitz-Hubsch T, Wullner U, Berthier E, Ollagnon-Roman E, Riess O, Forlani S, et al. Spinocerebellar ataxia type 11 (SCA11) is an uncommon cause of dominant ataxia among French and German kindreds. J. Neurol. Neurosurg. Psychiatry. 2010;81:1229–1232. - PubMed
-
- Houlden H. Spinocerebellar ataxia type 11. In: Pagon RA, Bird TD, Dolan CR, Stephens K, editors. GeneReviews. Seattle: University of Washington; 2008. PMID 20301723.
-
- Takahashi M, Tomizawa K, Sato K, Ohtake A, Omori A. A novel tau-tubulin kinase from bovine brain. FEBS Lett. 1995;372:59–64. - PubMed
-
- Manning G, Whyte DB, Martinez R, Hunter T, Sudarsanam S. The protein kinase complement of the human genome. Science. 2002;298:1912–1934. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases