

Breaking evolutionary and pleiotropic constraints in mammals: On sloths, manatees and homeotic mutations
- PMID: 21548920
- PMCID: PMC3120709
- DOI: 10.1186/2041-9139-2-11
Breaking evolutionary and pleiotropic constraints in mammals: On sloths, manatees and homeotic mutations
Erratum in
-
Correction to: Breaking evolutionary and pleiotropic constraints in mammals: on sloths, manatees and homeotic mutations.Evodevo. 2021 Nov 22;12(1):13. doi: 10.1186/s13227-021-00183-0. Evodevo. 2021. PMID: 34809711 Free PMC article. No abstract available.
Abstract
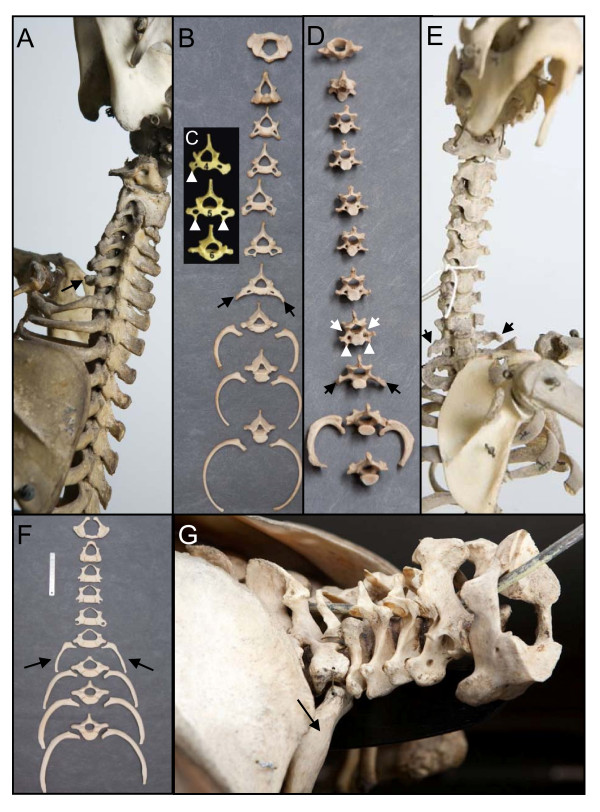
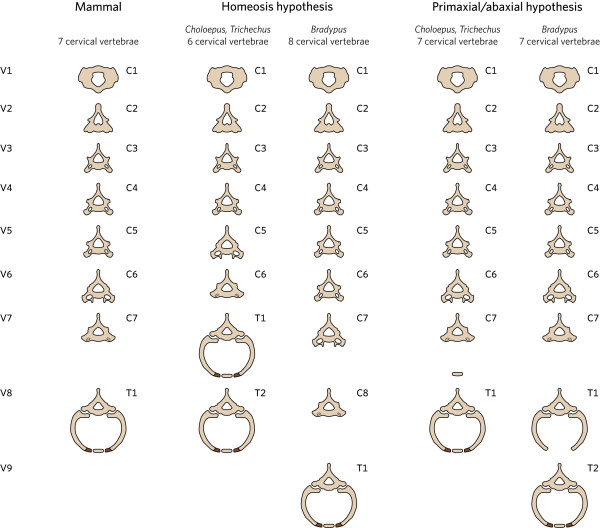
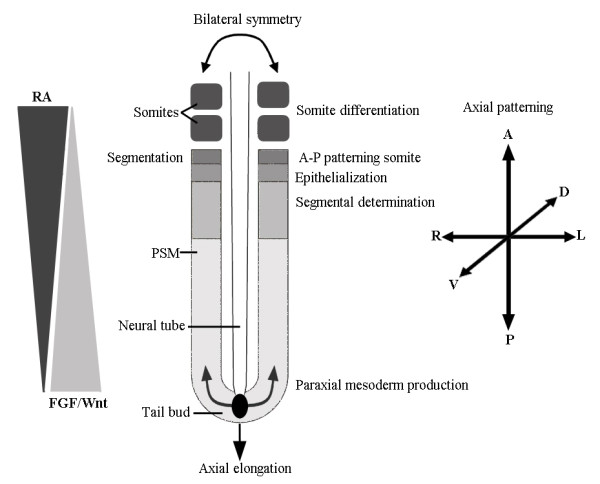
Background: Mammals as a rule have seven cervical vertebrae, except for sloths and manatees. Bateson proposed that the change in the number of cervical vertebrae in sloths is due to homeotic transformations. A recent hypothesis proposes that the number of cervical vertebrae in sloths is unchanged and that instead the derived pattern is due to abnormal primaxial/abaxial patterning.
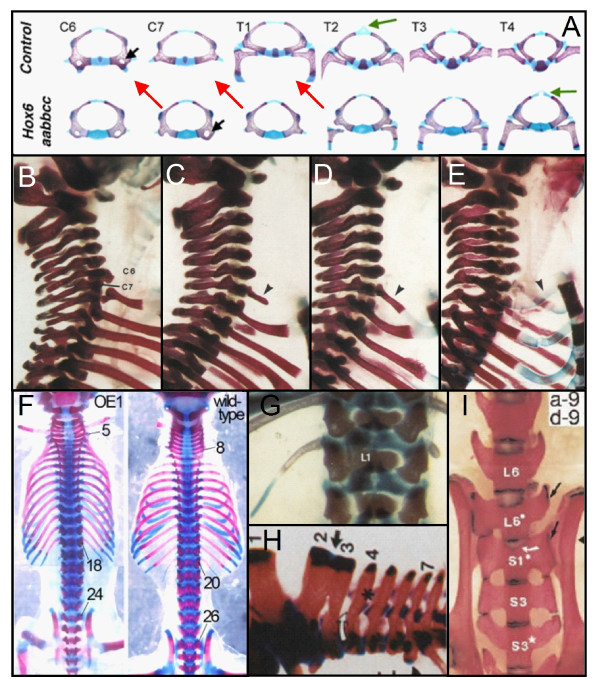
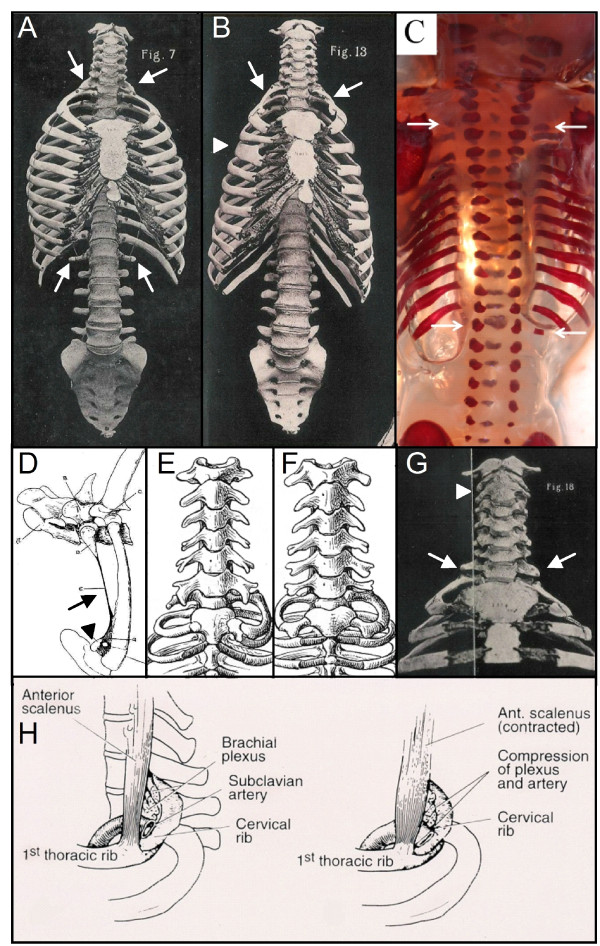
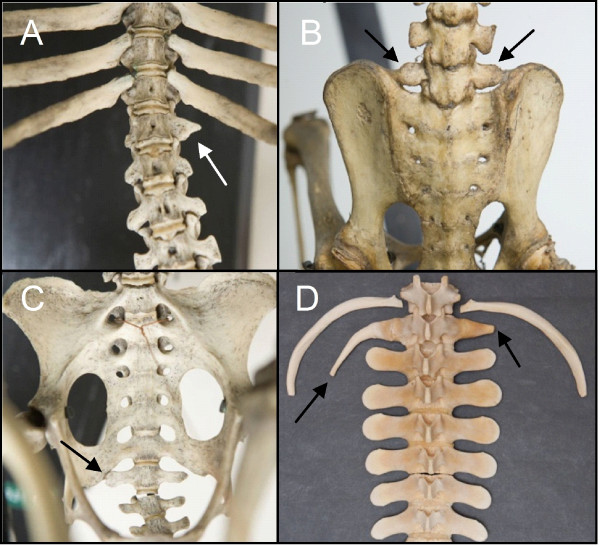
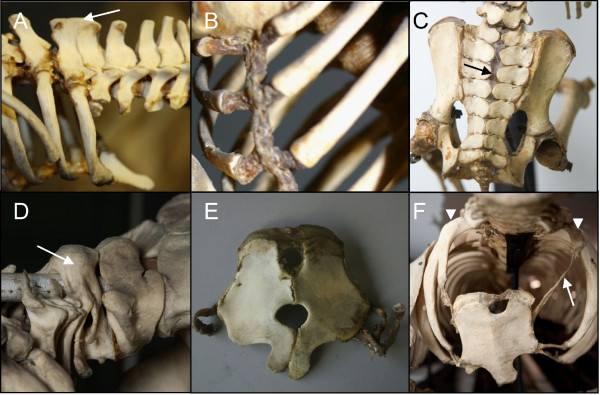
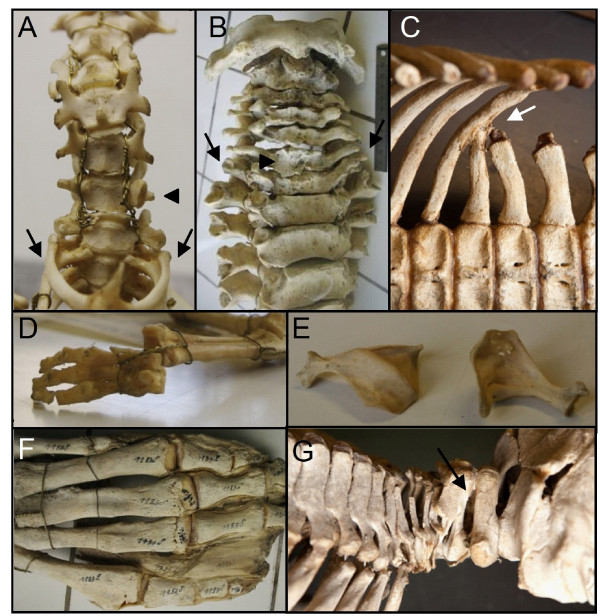
Results: We test the detailed predictions derived from both hypotheses for the skeletal patterns in sloths and manatees for both hypotheses. We find strong support for Bateson's homeosis hypothesis. The observed vertebral and rib patterns cannot be explained by changes in primaxial/abaxial patterning. Vertebral patterns in sloths and manatees are similar to those in mice and humans with abnormal numbers of cervical vertebrae: incomplete and asymmetric homeotic transformations are common and associated with skeletal abnormalities. In sloths the homeotic vertebral shift involves a large part of the vertebral column. As such, similarity is greatest with mice mutant for genes upstream of Hox.
Conclusions: We found no skeletal abnormalities in specimens of sister taxa with a normal number of cervical vertebrae. However, we always found such abnormalities in conspecifics with an abnormal number, as in many of the investigated dugongs. These findings strongly support the hypothesis that the evolutionary constraints on changes of the number of cervical vertebrae in mammals is due to deleterious pleitropic effects. We hypothesize that in sloths and manatees low metabolic and activity rates severely reduce the usual stabilizing selection, allowing the breaking of the pleiotropic constraints. This probably also applies to dugongs, although to a lesser extent.
Figures
References
-
- Colbert EU. Relationships of the Okapi. J Mammol. 1938;19:47–64. doi: 10.2307/1374281. - DOI
-
- Woolfenden GE. Postcranial morphology of the waterfowl. Bull Florida State Museum Biol Sci. 1961;6:1–129.
-
- Badlangana NL, Adams JW, Manger PR. The giraffe (Giraffa camelopardalis) cervical vertebral column: a heuristic example in understanding evolutionary processes? Zool J Linn Soc. 2009;155:736–757. doi: 10.1111/j.1096-3642.2008.00458.x. - DOI
-
- Leboucq H. Recherches sur les variations anatomiques de la première côte chez l'homme. Arch Biol. 1898;15:9–178.
-
- Schultz AH. In: Primatologia - Handbuch der Primatenkunde. Hofer H, Schultz AH, Starck D, editor. Basel/New York; 1961. Vertebral column and thorax; p. 66. Band 4, Lieferung 5.
LinkOut - more resources
Full Text Sources
Research Materials
