

Tyrosine phosphorylation of Munc18c on residue 521 abrogates binding to Syntaxin 4
- PMID: 21548926
- PMCID: PMC3103433
- DOI: 10.1186/1471-2091-12-19
Tyrosine phosphorylation of Munc18c on residue 521 abrogates binding to Syntaxin 4
Abstract
Background: Insulin stimulates exocytosis of GLUT4 from an intracellular store to the cell surface of fat and muscle cells. Fusion of GLUT4-containing vesicles with the plasma membrane requires the SNARE proteins Syntaxin 4, VAMP2 and the regulatory Sec1/Munc18 protein, Munc18c. Syntaxin 4 and Munc18c form a complex that is disrupted upon insulin treatment of adipocytes. Munc18c is tyrosine phosphorylated in response to insulin in these cells. Here, we directly test the hypothesis that tyrosine phosphorylation of Munc18c is responsible for the observed insulin-dependent abrogation of binding between Munc18c and Syntaxin 4.
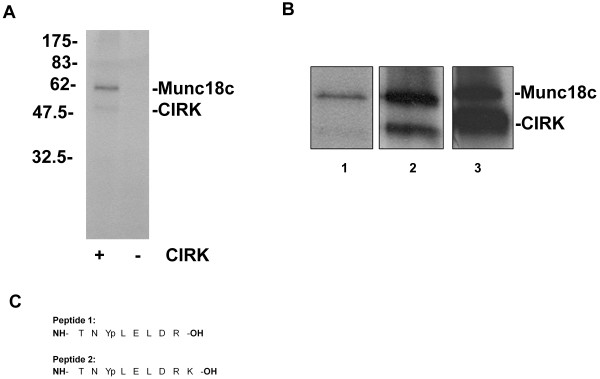
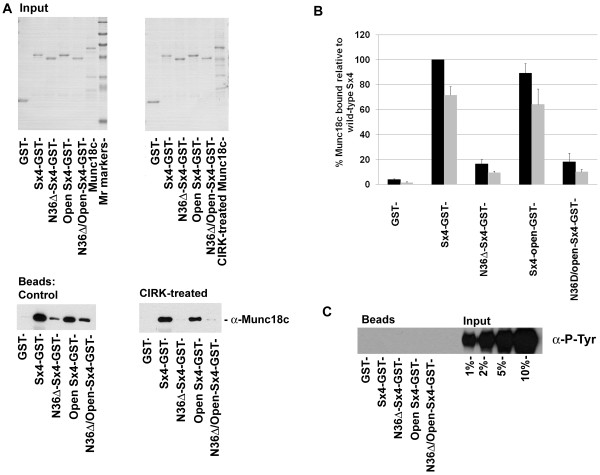
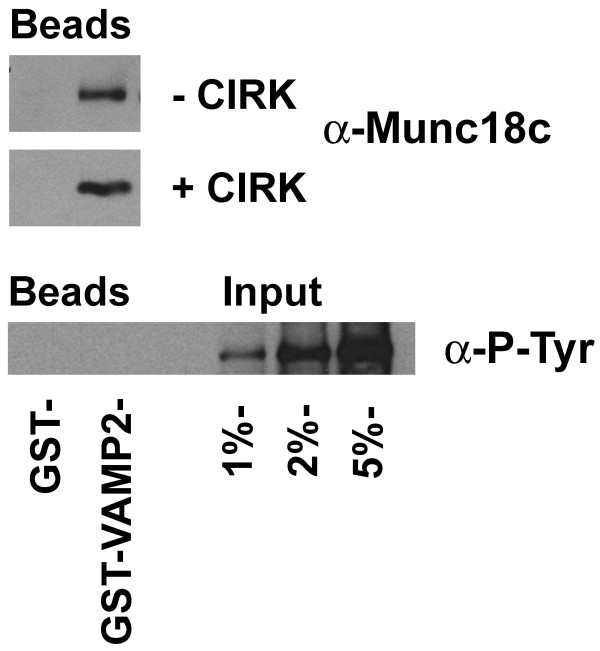
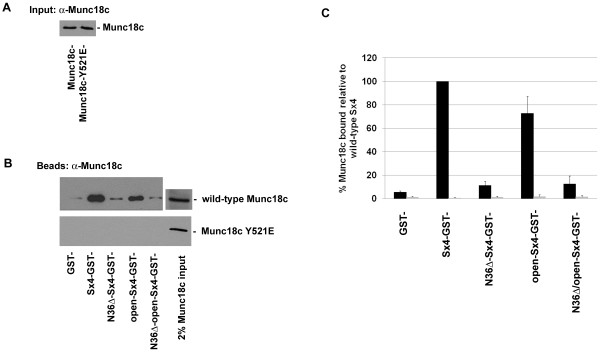
Results: We show that Munc18c is directly phosphorylated by recombinant insulin receptor tyrosine kinase in vitro. Using pull-down assays, we show that phosphorylation abrogates binding of Munc18c to both Syntaxin 4 and the v-SNARE VAMP2, as does the introduction of a phosphomimetic mutation into Munc18c (Y521E).
Conclusion: Our data indicate that insulin-stimulated tyrosine phosphorylation of Munc18c impairs the ability of Munc18c to bind its cognate SNARE proteins, and may therefore represent a regulatory step in GLUT4 traffic.
Figures
References
-
- Martin OJ, Lee A, McGraw TE. GLUT4 Distribution between the Plasma Membrane and the Intracellular Compartments Is Maintained by an Insulin-modulated Bipartite Dynamic Mechanism. J Biol Chem. 2006;281(1):484–490. - PubMed
-
- Hong W. SNAREs and traffic. Biochim Biophys Acta. 2005;1744(3):493–517. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
