

The requirement for the Dam1 complex is dependent upon the number of kinetochore proteins and microtubules
- PMID: 21549601
- PMCID: PMC3100407
- DOI: 10.1016/j.cub.2011.04.002
The requirement for the Dam1 complex is dependent upon the number of kinetochore proteins and microtubules
Abstract
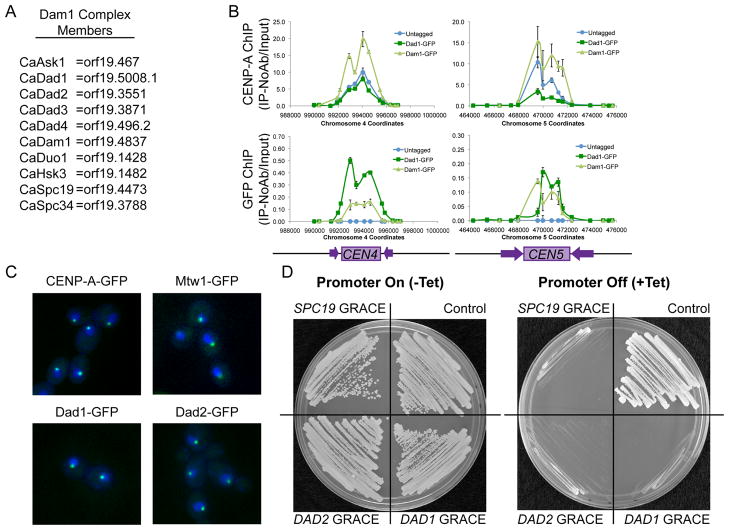
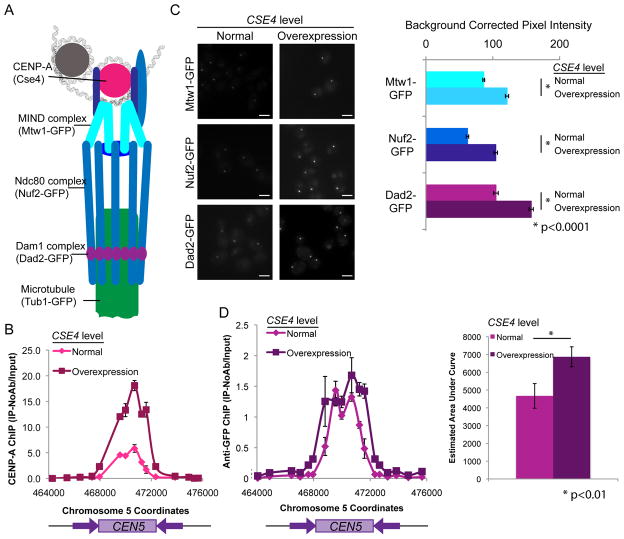
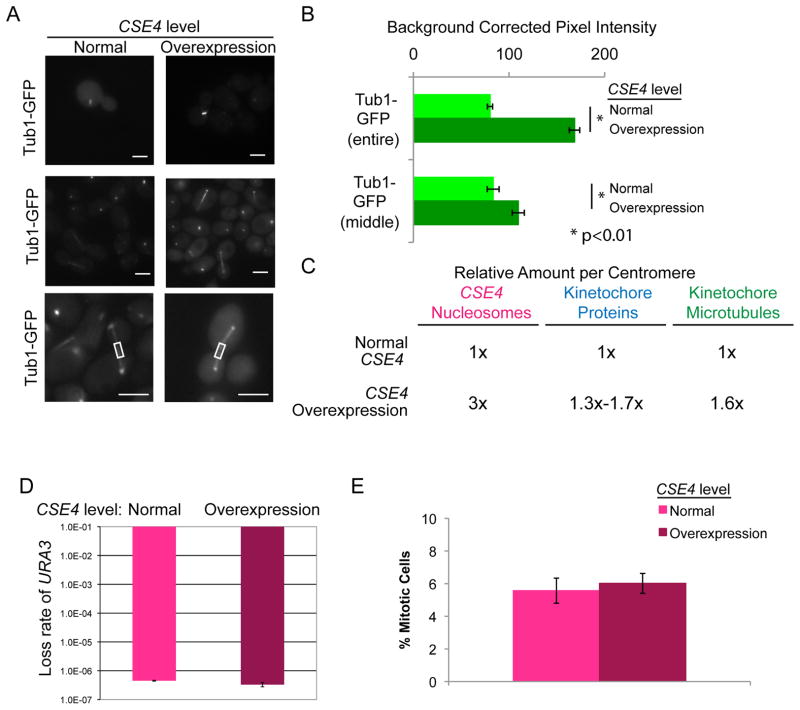
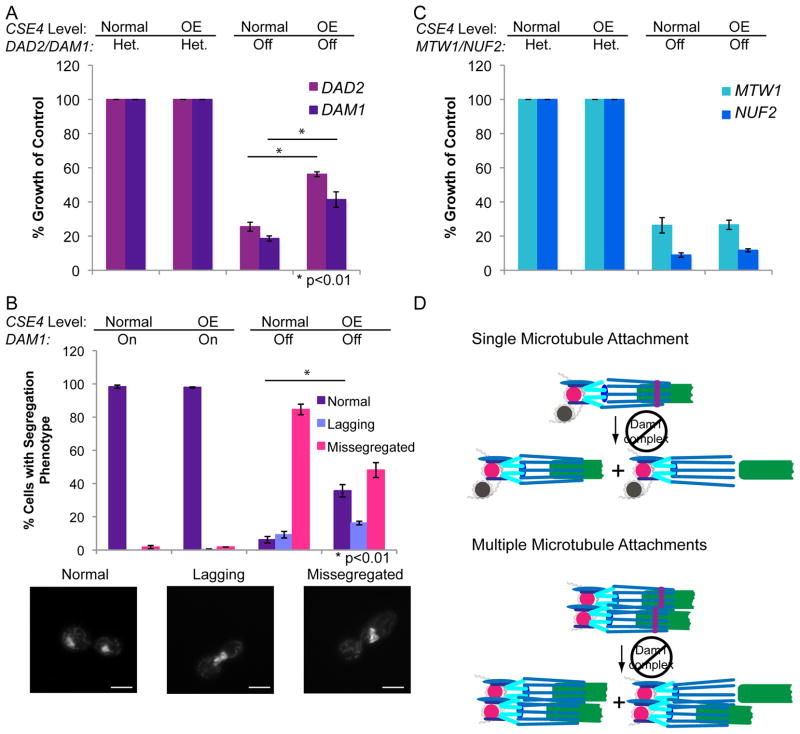
The Dam1 complex attaches the kinetochore to spindle microtubules and is a processivity factor in vitro. In Saccharomyces cerevisiae, which has point centromeres that attach to a single microtubule, deletion of any Dam1 complex member results in chromosome segregation failures and cell death. In Schizosaccharomyces pombe, which has epigenetically defined regional centromeres that each attach to 3-5 kinetochore microtubules, Dam1 complex homologs are not essential. To determine why the complex is essential in some organisms and not in others, we used Candida albicans, a multimorphic yeast with regional centromeres that attach to a single microtubule. Interestingly, the Dam1 complex was essential in C. albicans, suggesting that the number of microtubules per centromere is critical for its requirement. Importantly, by increasing CENP-A expression levels, more kinetochore proteins and microtubules were recruited to the centromeres, which remained fully functional. Furthermore, Dam1 complex members became less crucial for growth in cells with extra kinetochore proteins and microtubules. Thus, the requirement for the Dam1 complex is not due to the DNA-specific nature of point centromeres. Rather, the Dam1 complex is less critical when chromosomes have multiple kinetochore complexes and microtubules per centromere, implying that it functions as a processivity factor in vivo as well as in vitro.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
The essentiality of the fungus-specific Dam1 complex is correlated with a one-kinetochore-one-microtubule interaction present throughout the cell cycle, independent of the nature of a centromere.Eukaryot Cell. 2011 Oct;10(10):1295-305. doi: 10.1128/EC.05093-11. Epub 2011 May 13. Eukaryot Cell. 2011. PMID: 21571923 Free PMC article.
-
Outer kinetochore protein Dam1 promotes centromere clustering in parallel with Slk19 in budding yeast.Chromosoma. 2019 Jun;128(2):133-148. doi: 10.1007/s00412-019-00694-9. Epub 2019 Mar 12. Chromosoma. 2019. PMID: 30903360
-
Molecular architecture of the kinetochore-microtubule attachment site is conserved between point and regional centromeres.J Cell Biol. 2008 May 19;181(4):587-94. doi: 10.1083/jcb.200803027. Epub 2008 May 12. J Cell Biol. 2008. PMID: 18474626 Free PMC article.
-
Dam1 is the right one: phosphoregulation of kinetochore biorientation.Dev Cell. 2002 Nov;3(5):610-1. doi: 10.1016/s1534-5807(02)00332-5. Dev Cell. 2002. PMID: 12431367 Review.
-
Family matters: structural and functional conservation of centromere-associated proteins from yeast to humans.Trends Cell Biol. 2013 Jun;23(6):260-9. doi: 10.1016/j.tcb.2013.01.010. Epub 2013 Mar 5. Trends Cell Biol. 2013. PMID: 23481674 Review.
Cited by
-
A Matter of Scale and Dimensions: Chromatin of Chromosome Landmarks in the Fungi.Microbiol Spectr. 2017 Jul;5(4):10.1128/microbiolspec.funk-0054-2017. doi: 10.1128/microbiolspec.FUNK-0054-2017. Microbiol Spectr. 2017. PMID: 28752814 Free PMC article. Review.
-
Diversity in requirement of genetic and epigenetic factors for centromere function in fungi.Eukaryot Cell. 2011 Nov;10(11):1384-95. doi: 10.1128/EC.05165-11. Epub 2011 Sep 9. Eukaryot Cell. 2011. PMID: 21908596 Free PMC article. Review.
-
Gene Essentiality Analyzed by In Vivo Transposon Mutagenesis and Machine Learning in a Stable Haploid Isolate of Candida albicans.mBio. 2018 Oct 30;9(5):e02048-18. doi: 10.1128/mBio.02048-18. mBio. 2018. PMID: 30377286 Free PMC article.
-
CaMad2 Promotes Multiple Aspects of Genome Stability Beyond Its Direct Function in Chromosome Segregation.Genes (Basel). 2019 Dec 5;10(12):1013. doi: 10.3390/genes10121013. Genes (Basel). 2019. PMID: 31817479 Free PMC article.
-
Structural organization of very small chromosomes: study on a single-celled evolutionary distant eukaryote Giardia intestinalis.Chromosoma. 2015 Mar;124(1):81-94. doi: 10.1007/s00412-014-0486-5. Epub 2014 Aug 30. Chromosoma. 2015. PMID: 25171919
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
