

Peptide signaling in plant development
- PMID: 21549958
- PMCID: PMC3139689
- DOI: 10.1016/j.cub.2011.03.012
Peptide signaling in plant development
Abstract
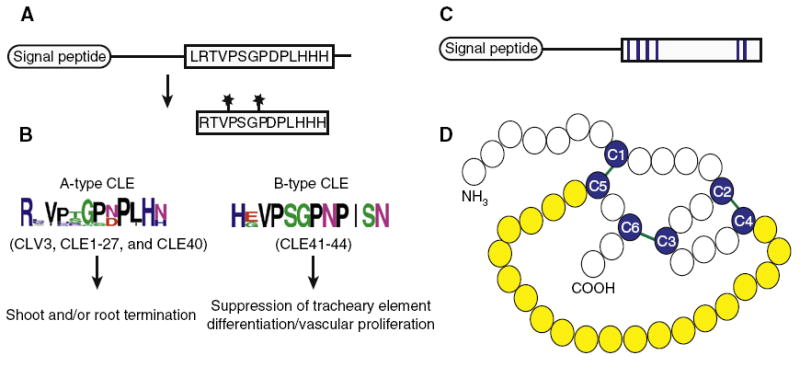
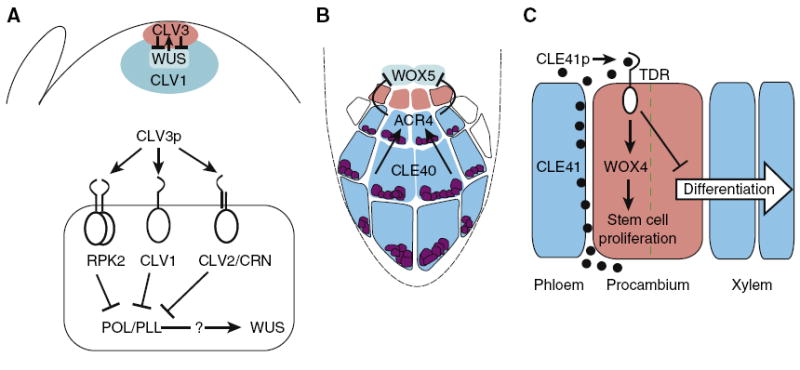
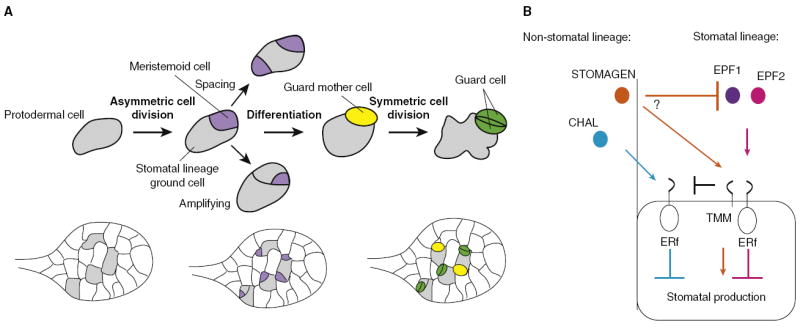
Cell-to-cell communication is integral to the evolution of multicellularity. In plant development, peptide signals relay information coordinating cell proliferation and differentiation. These peptides are often encoded by gene families and bind to corresponding families of receptors. The precise spatiotemporal expression of signals and their cognate receptors underlies developmental patterning, and expressional and biochemical changes over evolutionary time have likely contributed to the refinement and complexity of developmental programs. Here, we discuss two major plant peptide families which have central roles in plant development: the CLAVATA3/ENDOSPERM SURROUNDING REGION (CLE) peptide family and the EPIDERMAL PATTERNING FACTOR (EPF) family. We discuss how specialization has enabled the CLE peptides to modulate stem cell differentiation in various tissue types, and how differing activities of EPF peptides precisely regulate the stomatal developmental program, and we examine the contributions of these peptide families to plant development from an evolutionary perspective.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures

References
-
- Dohlman HG, Thorner JW. Regulation of G protein-initiated signal transduction in yeast: paradigms and principles. Annu Rev Biochem. 2001;70:703–754. - PubMed
-
- Takayama S, Isogai A. Self-incompatibility in plants. Annu Rev Plant Biol. 2005;56:467–489. - PubMed
-
- Butenko MA, Vie AK, Brembu T, Aalen RB, Bones AM. Plant peptides in signalling: looking for new partners. Trends Plant Sci. 2009;14:255–263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
