

Review
doi: 10.1016/B978-0-12-387036-0.00005-0.
Zebrafish lipid metabolism: from mediating early patterning to the metabolism of dietary fat and cholesterol
Affiliations
- PMID: 21550441
- PMCID: PMC3593232
- DOI: 10.1016/B978-0-12-387036-0.00005-0
Item in Clipboard
Review
Zebrafish lipid metabolism: from mediating early patterning to the metabolism of dietary fat and cholesterol
Methods Cell Biol.
2011.
Abstract
Lipids serve essential functions in cells as signaling molecules, membrane components, and sources of energy. Defects in lipid metabolism are implicated in a number of pandemic human diseases, including diabetes, obesity, and hypercholesterolemia. Many aspects of how fatty acids and cholesterol are absorbed and processed by intestinal cells remain unclear and present a hurdle to developing approaches for disease prevention and treatment. Numerous studies have shown that the zebrafish is an excellent model for vertebrate lipid metabolism. In this chapter, we review studies that employ zebrafish to better understand lipid signaling and metabolism.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
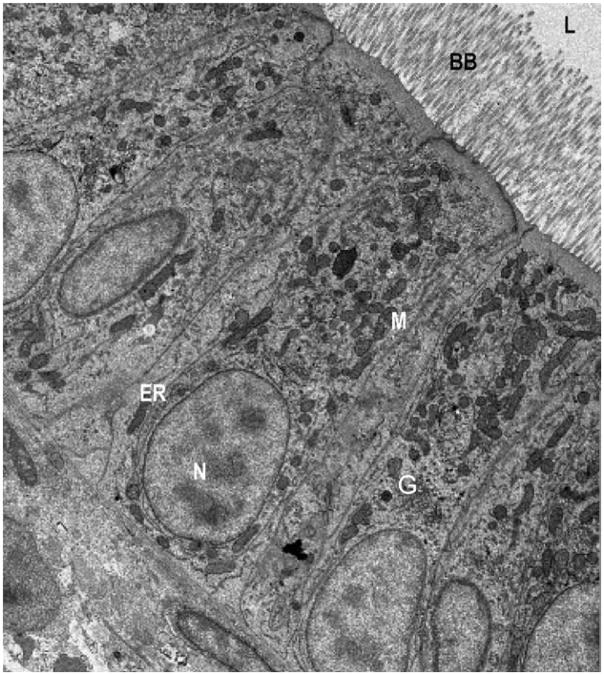
The intestinal enterocytes of zebrafish and mammals exhibit a high degree of morphological similarity. Electron micrograph of enterocytes from a larval zebrafish (6 dpf). Zebrafish enterocytes exhibit the typical characteristics of mammalian polarized intestinal cells including apical microvilli, which extend into the intestinal lumen (L) and form the brush border, as well as basal nuclei (N). Organelles and subcellular details including mitochondria (M), Golgi bodies (G), and endoplasmic reticulum (ER) are apparent throughout the enterocytes. For comparison, see mouse EMs in Buhman et al. (2002).
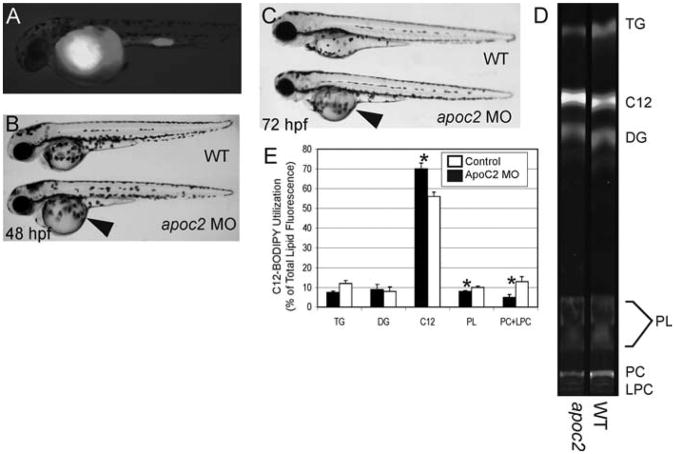
BODIPY lipid analogs enable studies of yolk metabolism during early zebrafish development. To assay the function of apoc2 during zebrafish development, embryos were injected with an apoc2 morpholino at the 1–4 cell stage followed by injection of a fluorescent fatty acid (BODIPY-C12) at 24 h postfertilization (hpf). (A) At 48 hpf, embryos injected with BODIPY-C12 retain the fluorescent analog primarily in their yolk. (B, C) apoc2 morphants exhibit an enlarged yolk phenotype (arrowhead) at 48 and 72 hpf, indicating that apoc2 is necessary for yolk utilization during larval development. (D, E) To determine the metabolic consequences of apoc2 deficiency, 1 dpf wild-type and apoc2 morphant larvae were injected with BODIPY-C12 and assayed using fluorescent thin layer chromatography (TLC) 2 days later. (D) TLC analysis shows that BODIPY-C12 is incorporated primarily into triacylglycerol (TAG), diacylglycerol (DG), and phosphatidylcholine (PC) in both wild-type and apoc2 morphant larvae. (E) apoc2 morphants exhibit defects in PC and lysophosphatidylcholine (LPC) metabolism. Total lipid fluorescence was quantified from TLC plates run with total lipids extracted from wild-type and apoc2 morphants. Triacylglycerol (TAG), diacylglycerol (DG), BODIPY C12:0 (C12), phospholipids (PL), PC, and LPC.* p < 0.05.
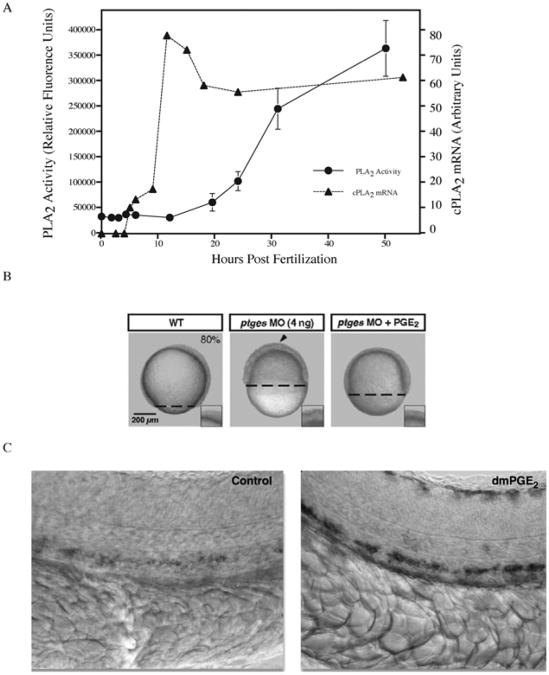
Phospholipase activity is required for proper zebrafish development. (A) Cytosolic phospholipase A2 (cPLA2) mRNA levels and PLA2 enzymatic activity were quantified from whole embryo lysates during early stages of zebrafish development. cPLA2 expression peaks during somitogenesis (10 h) while enzymatic activity steadily increases as development proceeds. PLA2 activity is required to generate prostaglandins. (B) Blocking prostaglandin production during early zebrafish development through inhibition of prostaglandin–endoperoxide synthase (Ptgs1) via morpholino knockdown results in developmental arrest at epiboly. Developmental arrest can be rescued by adding back the exogenous enzyme product (PGE2). Reproduced with permission by Development (Speirs et al., 2010). (C) Embryos exposed to the PGE2 analog (16,16-dimethyl-PGE2; dmPGE2) exhibit increased expression of runx11 and cmyb1 as evidenced by in situ hybridization. These genes are expressed in the ventral wall of the dorsal aorta in a region analogous to the mammalian aorta–gonad–mesonephros and are required for mammalian hematopoietic stem cell development. Reprinted by permission from Macmillan Publishers Ltd.: Nature (North et al., 2007). Prostaglandin E2 regulates vertebrate haematopoietic stem cell homeostasis. Nature 447, 1007–11, copyright 2007.
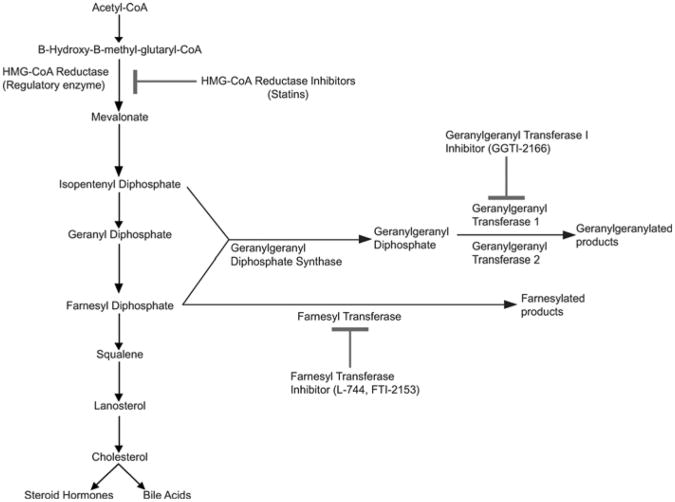
Cholesterol biosynthesis and protein lipidation are highly conserved in vertebrates. Due to the high genetic conservation of these pathways, human drugs can be used to block specific biochemical steps to determine the roles that cholesterol and protein lipidation play in zebrafish development. Reprinted from Development Cell with permission from Elsevier (Thorpe et al., 2004).

Embryos treated with statins exhibit developmental defects. In comparison to untreated embryos (A), embryos soaked in a low dose of mevinolin (0.06 μM). (B) exhibit mild developmental defects, as evidenced by tail kinks. (C) Exposure to higher doses of mevinolin (1.2 μM) results in blunted axis elongation, necrosis and developmental arrest. (D) Simvastatin treated embryos (2.0 μM) exhibit similar developmental defects. (E) Dose response of mevinolin on developmental arrest (mean + SEM, n = 3). (F) Dose response of simvastatin on developmental arrest (mean + SEM, n = 3). Reprinted from Development Cell with permission from Elsevier (Thorpe et al., 2004).
Isoprenoid intermediates rescue the developmental defects caused by statin treatment. Embryos at early cell stages were injected with mevalonate and then soaked overnight in mevinolin, simvastatin, or atorvastatin. (A) Embryos treated with mevinolin show severe developmental defects at 24 hpf. (B) Embryos injected with the isoprenoid intermediate mevalonate (1–16 cell stages) and then treated with mevinolin exhibit normal morphology. (C) Mevalonate injection rescues the somatic defects observed in embryos treated with different statins. Embryo morphology was scored at 24 hpf. Data represent the MEAN ± SEM from 3–4 experiments. Reprinted from Development Cell with permission from Elsevier (Thorpe et al., 2004).
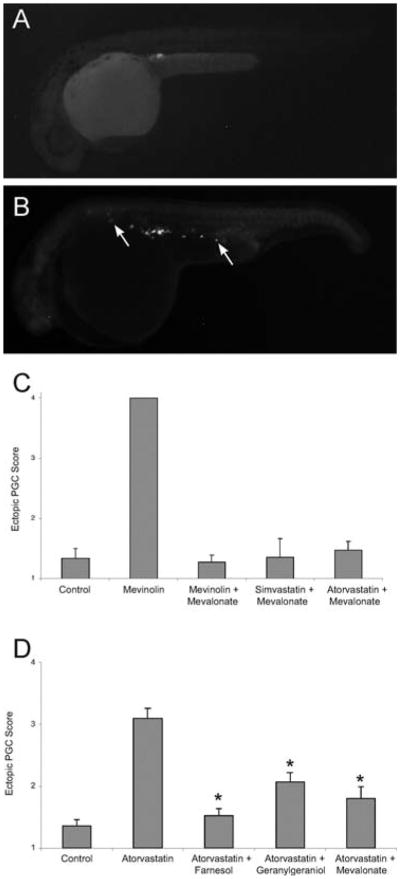
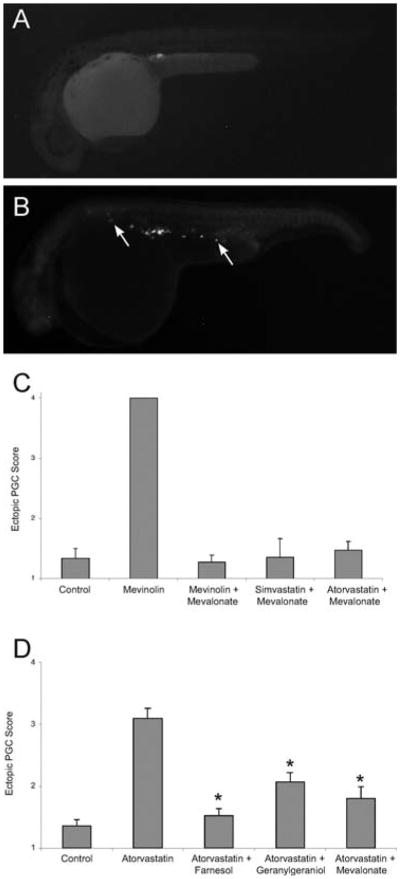
Statin treatment causes abnormal primordial germ cell (PGC) migration in zebrafish embryos. Compared to wild-type (A), embryos treated with statins (B) display ectopic PGCs. The arrows indicate ectopic PGCs that have failed to migrate to the developing gonad. (C) The PGC migratory defect observed following statin treatment is prevented by injections of isoprenoid intermediates. Embryos at early stages (1–16 cell) were injected with gfp-nos mRNA and mevalonate and then soaked overnight in mevinolin, simvastatin or atorvastatin. At 24 hpf, embryos were scored for ectopic PGCs, with a score of 1 indicating a wild-type single gonadal cluster and score of 4 indicating no discernable PGC cluster. Data represent the mean ± SEM from 3–4 experiments. (D) The PGC migratory defect observed following statin treatment is prevented by increasing the levels of isoprenoid synthesis intermediates. Embryos injected at the 1–16 cell stage with farnesol, geranylgeraniol or mevalonate and then soaked overnight with atorvastatin (10 μM) show normal PGC migration. Data represents MEAN ± SEM, *p < 0.01 difference from atorvastatin alone. Reprinted from Development Cell with permission from Elsevier (Thorpe et al., 2004).
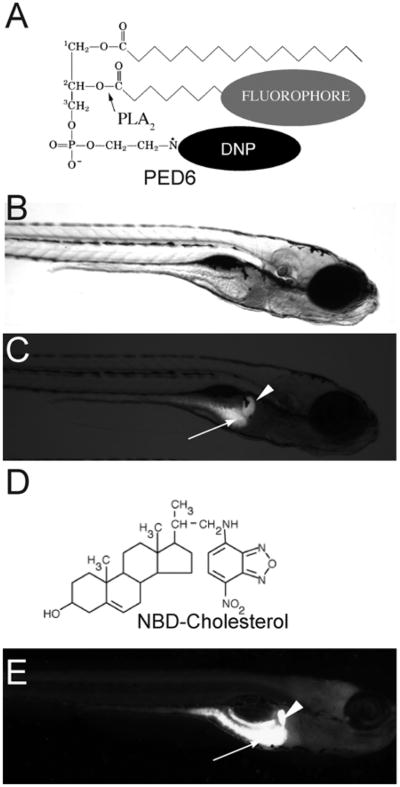
PED6 and NBD-cholesterol visualize lipid uptake in larval zebrafish. (A) The chemical structures of PED6. The BODIPY-labeled acyl chain of PED6 is normally quenched by the dinitrophenyl group at the sn-3 position. Upon PLA2 cleavage at the sn-2 position, the BODIPY-labeled acyl chain is unquenched and can fluoresce. Bright field (B) and fluorescent (C) images of 5 dpf larva following soaking in PED6 for 6 h. PED6 labeling reveals lipid processing in the gall bladder (arrowhead) and intestine (arrow). (D) The chemical structure of NBD-cholesterol. The NBD-cholesterol analog contains a NDB fluorophore where the alkyl tail at the terminal end of cholesterol would normally reside. (E) Soaking zebrafish larvae (5 dpf) in NBD-cholesterol (3 mg/ml, solubilized with fish bile) for 2 h visualizes cholesterol uptake in the gall bladder (arrowhead) and intestine (arrow).
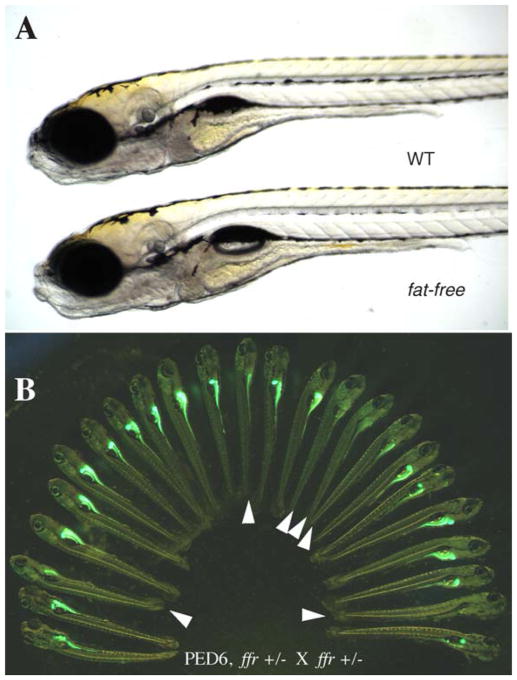
PED6 labeling of fat-free(ffr) larvae reveals defective lipid processing. (A) ffr larvae appear morphologically normal in comparison to wild-type siblings (6 dpf). (B). ffr mutants have diminished intestinal and gall bladder fluorescence when labeled with PED6, indicating abnormal lipid processing.
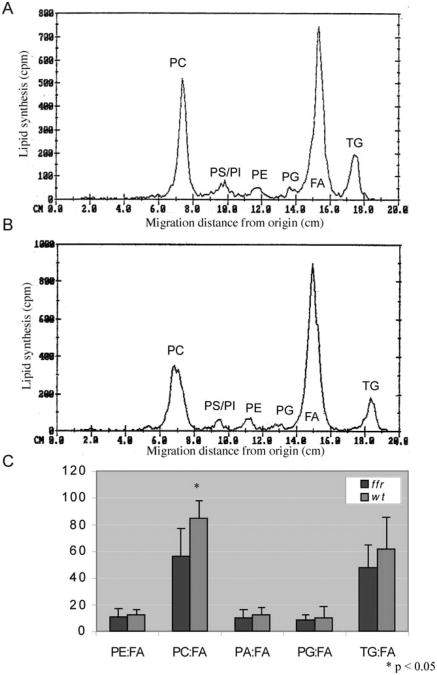
ffr larvae have abnormal phospholipid metabolism as shown by thin layer chromatography (TLC). To identify the metabolic defects in ffr larvae, mutants and wild-type larvae (4 dpf) were incubated with radioactive oleic acid (C18:1) for 20 h, followed by total lipid extraction and radioactive TLC analysis. (A, B) Chromatograms generated from scans of TLC plates run with total lipids extracted from wild-type (A) and ffr (B) larvae. They-axis reflects radioactivity counts (cpm; 2–3% of actual activity detected for 3H); the x-axis shows various oleic acid metabolites determined by the migration distance from the start of the TLC plate measured in centimeters (cm). The major metabolites derived from oleic acid (here FA) are PC, phosphatidylserine (PS), phosphatidylinositol (PI), phosphatidylethanolamine (PE), phosphatidylglycerol (PG), and TAG. (C) Comparison of relative FA metabolites between ffr and wild-type larvae reveals that ffr mutants have significantly decreased PC production (n = 9, mean ± SD). Reprinted from Methods in Enzymology with permission from Elsevier (Ho et al., 2003).
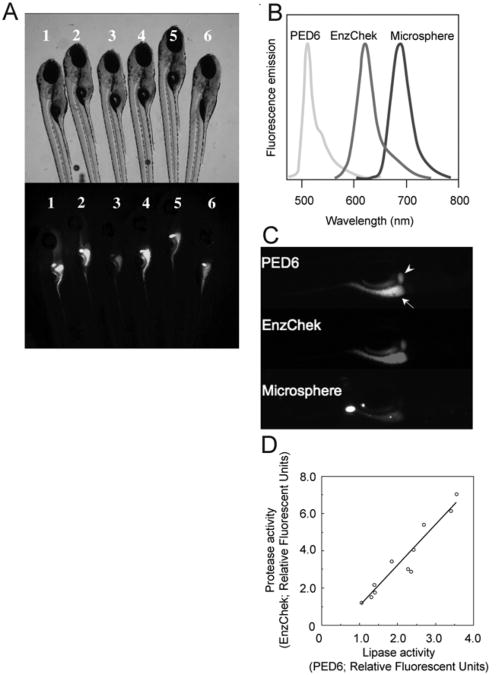
Concurrent feeding of PED6, EnzChek, and fluorescent microspheres enables the assessment of larval digestive function. To correct for interindividual variability observed in wild-type siblings (6 dpf) labeled with PED6 (A), multiple reporters of the digestive function can be used. (B) PED6, EnzChek and microspheres each fluoresce at distinct wavelengths, allowing simultaneous screening for lipase and protease activities, and swallowing function, respectively. (C) PED6 and EnzChek signal in the gall bladder (arrowhead) and intestine (arrow) of wild-type zebrafish. Microspheres in the intestine indicate normal ingestion. (D) Intestinal protease and phospholipase activity correlate, validating the use of the ratio of PED6 to EnzChek signal as readout of the digestive function. Figure from Hama et al. (2008), Am. J. Physiol. Gastrointest. Liver Physiol., used with permission from Am. Physiol. Soc.
References
-
- Apelqvist A, Li H, Sommer L, Beatus P, Anderson DJ, Honjo T, Hrabe de Angelis M, Lendahl U, Edlund H. Notch signalling controls pancreatic cell differentiation. Nature. 1999;400:877–881. - PubMed
-
- Begle A, Tryoen-Toth P, de Barry J, Bader MF, Vitale N. ARF6 regulates the synthesis of fusogenic lipids for calcium-regulated exocytosis in neuroendocrine cells. J Biol Chem. 2009;284:4836–4845. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
