

Nonsense-mediated mRNA decay (NMD) in animal embryogenesis: to die or not to die, that is the question
- PMID: 21550797
- PMCID: PMC3150509
- DOI: 10.1016/j.gde.2011.03.008
Nonsense-mediated mRNA decay (NMD) in animal embryogenesis: to die or not to die, that is the question
Abstract
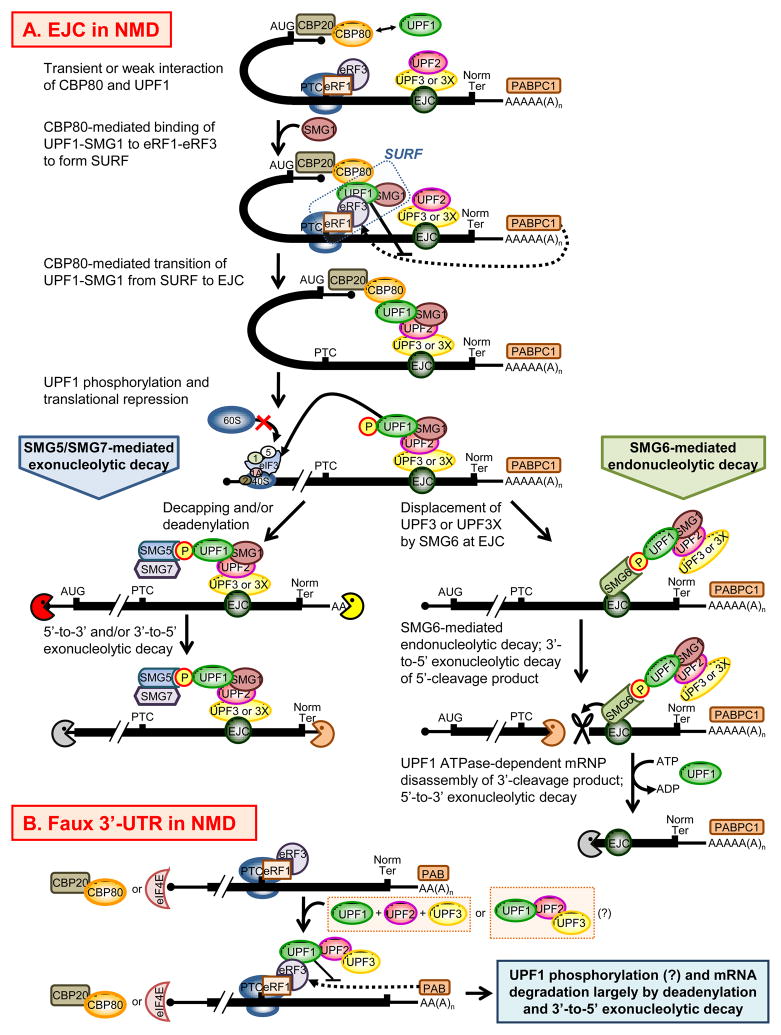
Nonsense-mediated mRNA decay (NMD) is a well-studied cellular quality-control pathway. It decreases the half-lives of eukaryotic mRNAs that aberrantly contain premature termination codons and additionally regulates an estimated 10-20% of normal transcripts. NMD factors play crucial roles during embryogenesis in many animals. Here, we review data indicating that NMD factors are required for proper embryogenesis by discussing the abnormal developmental phenotypes that result when the abundance of individual NMD factors is either downregulated or completely eliminated. We conclude that while NMD per se affects the embryogenesis of all animals, it is required to avoid embryonic lethality in only some animals. The critical roles of many NMD factors in other metabolic pathways undoubtedly also contribute to embryonic development if not viability.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Bhuvanagiri M, Schlitter AM, Hentze MW, Kulozik AE. NMD: RNA biology meets human genetic medicine. Biochem J. 2010;430:365–377. - PubMed
-
- Nicholson P, Mühlemann O. Cutting the nonsense: the degradation of PTC-containing mRNAs. Biochem Soc Trans. 2010;38:1615–1620. - PubMed
-
- Jaillon O, Bouhouche K, Gout JF, Aury JM, Noel B, Saudemont B, Nowacki M, Serrano V, Porcel BM, Segurens B, et al. Translational control of intron splicing in eukaryotes. Nature. 2008;451:359–362. - PubMed
-
- Mitrovich QM, Anderson P. mRNA surveillance of expressed pseudogenes in C. elegans. Curr Biol. 2005;15:963–967. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
