

The LRRK2-related Roco kinase Roco2 is regulated by Rab1A and controls the actin cytoskeleton
- PMID: 21551065
- PMCID: PMC3128523
- DOI: 10.1091/mbc.E10-12-0937
The LRRK2-related Roco kinase Roco2 is regulated by Rab1A and controls the actin cytoskeleton
Abstract
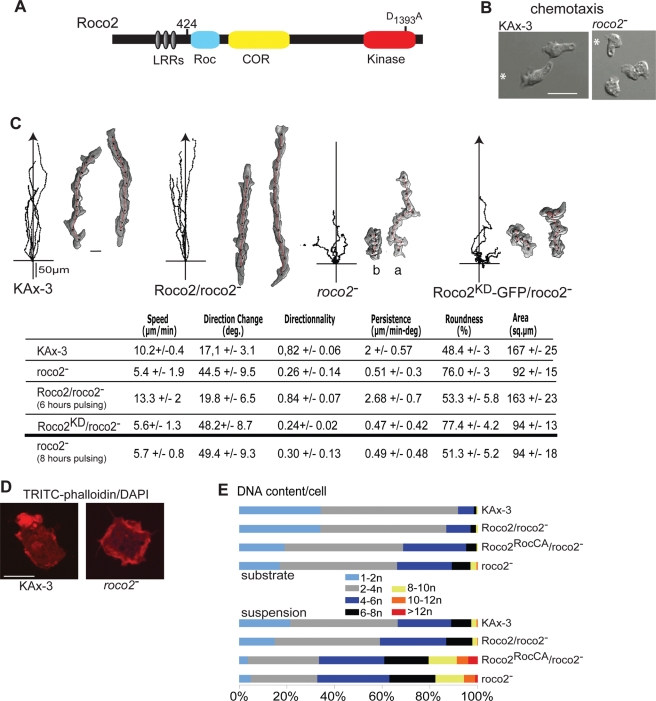
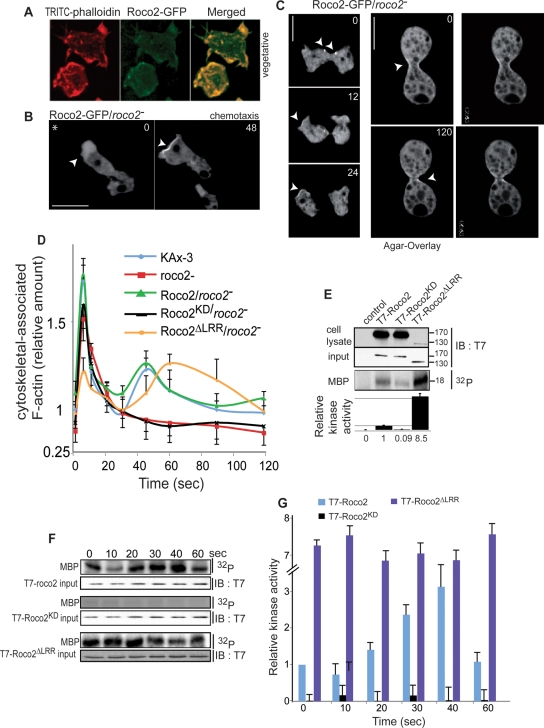
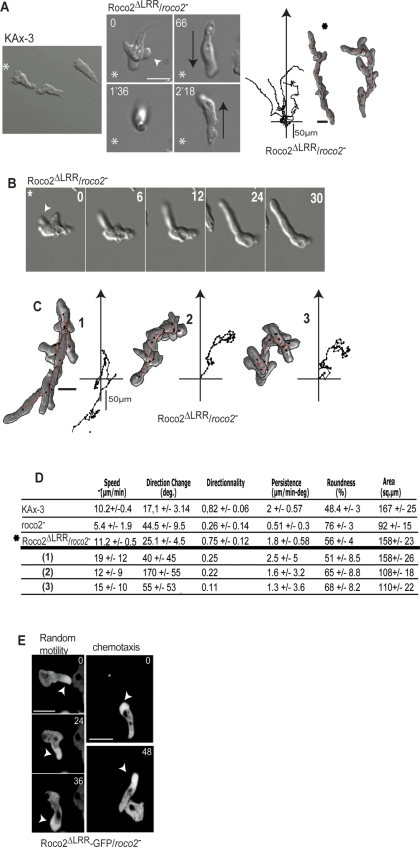
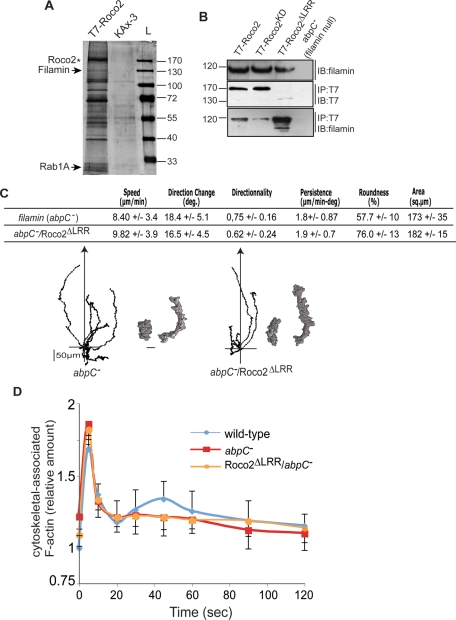
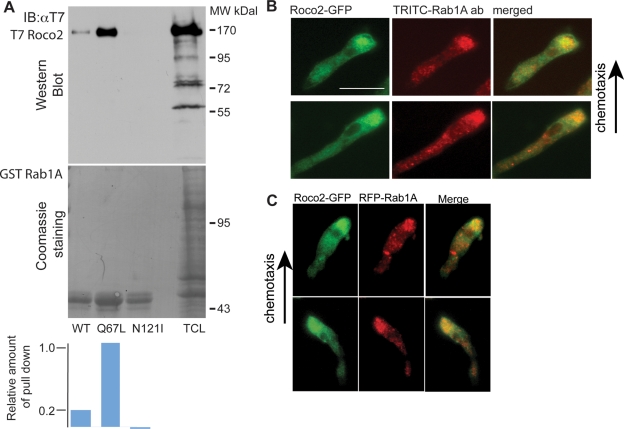
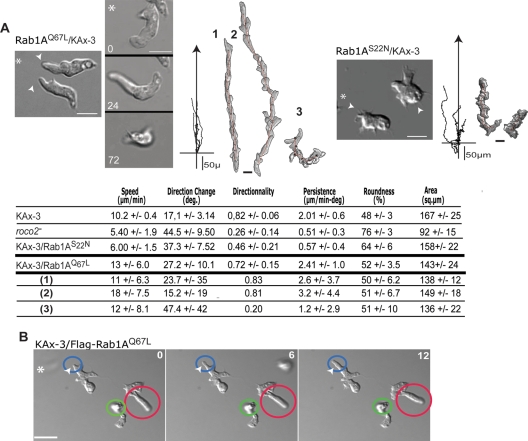
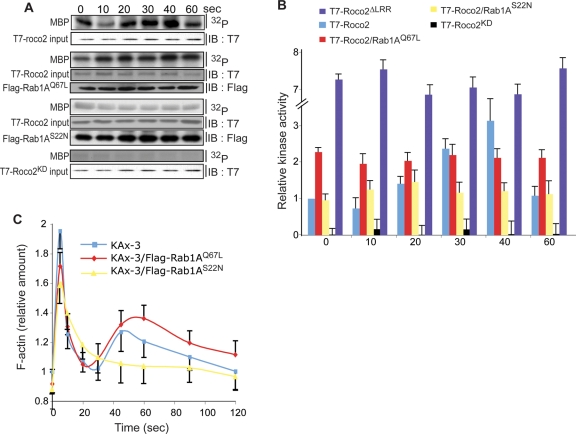
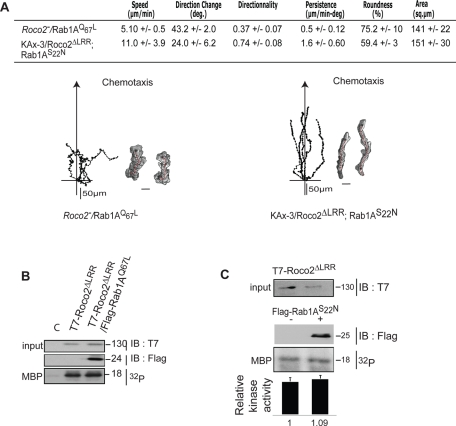
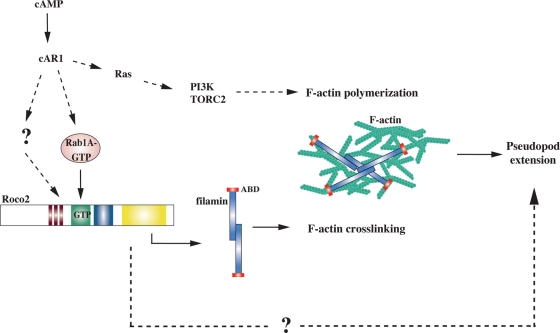
We identify a new pathway that is required for proper pseudopod formation. We show that Roco2, a leucine-rich repeat kinase 2 (LRRK2)-related Roco kinase, is activated in response to chemoattractant stimulation and helps mediate cell polarization and chemotaxis by regulating cortical F-actin polymerization and pseudopod extension in a pathway that requires Rab1A. We found that Roco2 binds the small GTPase Rab1A as well as the F-actin cross-linking protein filamin (actin-binding protein 120, abp120) in vivo. We show that active Rab1A (Rab1A-GTP) is required for and regulates Roco2 kinase activity in vivo and that filamin lies downstream from Roco2 and controls pseudopod extension during chemotaxis and random cell motility. Therefore our study uncovered a new signaling pathway that involves Rab1A and controls the actin cytoskeleton and pseudopod extension, and thereby, cell polarity and motility. These findings also may have implications in the regulation of other Roco kinases, including possibly LRRK2, in metazoans.
Figures
References
-
- Bandala-Sanchez E, Annesley SJ, Fisher PR. A phototaxis signalling complex in Dictyostelium discoideum. Eur J Cell Biol. 2006;85:1099–1106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
