

Gene loss and parallel evolution contribute to species difference in flower color
- PMID: 21551271
- PMCID: PMC3203625
- DOI: 10.1093/molbev/msr109
Gene loss and parallel evolution contribute to species difference in flower color
Abstract
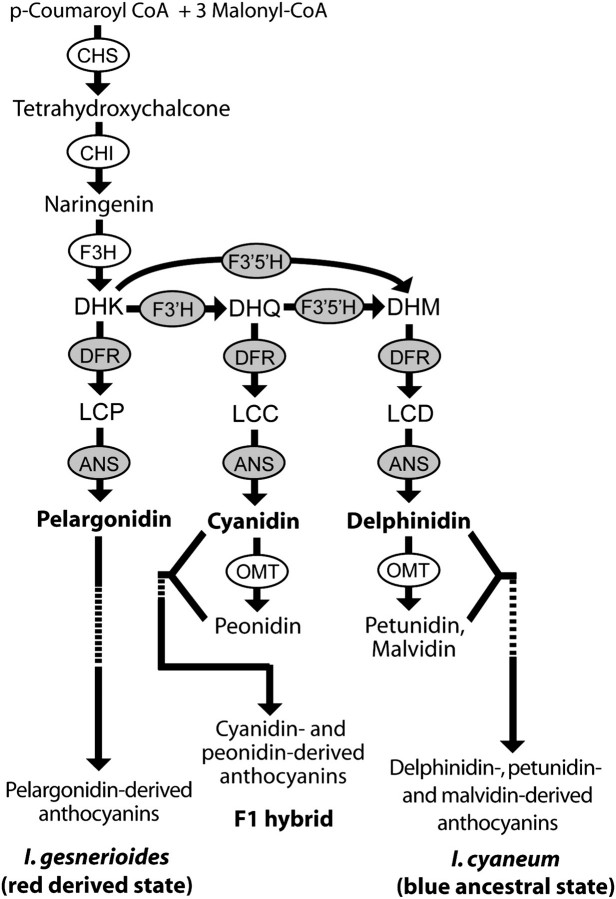
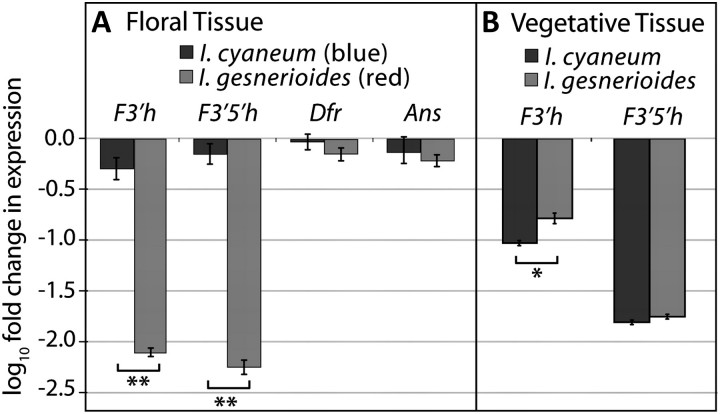
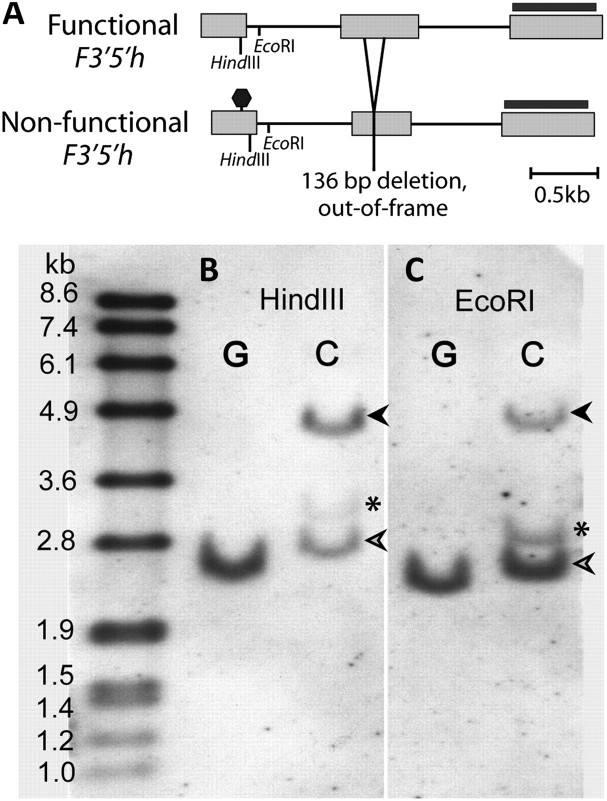
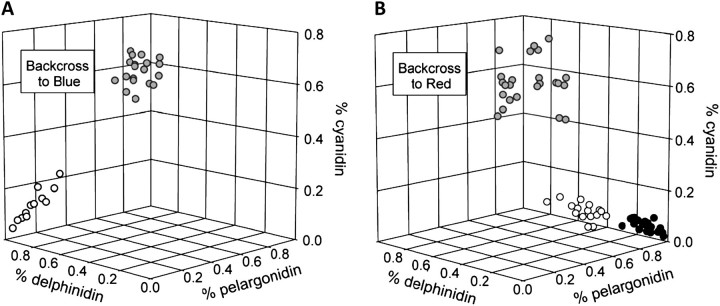
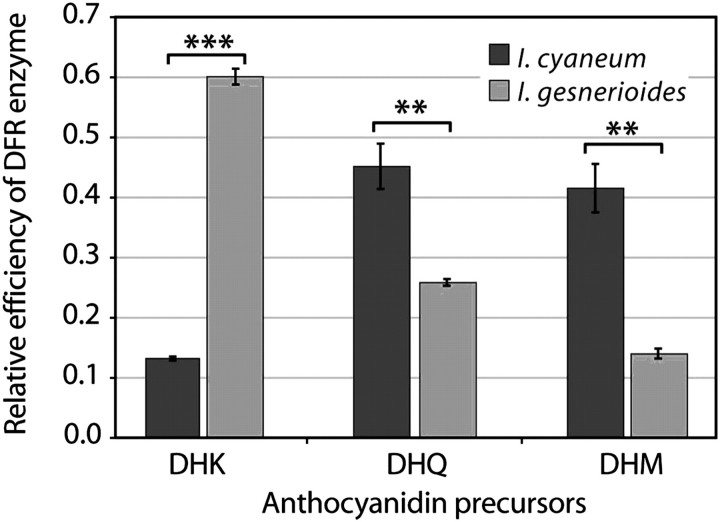
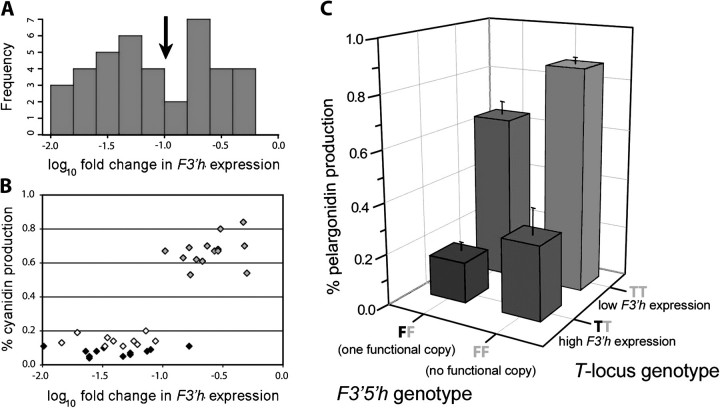
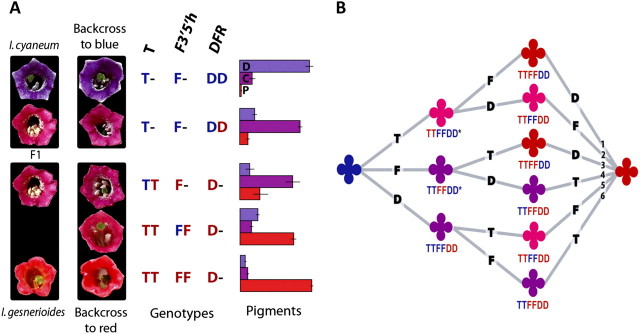
Although the importance of regulatory and functional sequence evolution in generating species differences has been studied to some extent, much less is known about the role of other types of genomic changes, such as fluctuation in gene copy number. Here, we apply analyses of gene function and expression of anthocyanin pigment pathway genes, as well as cosegregation analyses in backcross populations, to examine the genetic changes involved in the shift from blue to red flowers in Andean Iochroma (Solanaceae). We demonstrate that deletion of a gene coding for an anthocyanin pathway enzyme was necessary for the transition to red floral pigmentation. The downregulation of a second pathway gene was also necessary for the novel flower color, and this regulatory pattern parallels the genetic change in the two other red-flowered species in the sister family Convolvulaceae in which flower color change has been examined genetically. Finally, we document a shift in enzymatic function at a third locus, but the importance of this change in the transition to red flowers depends on the exact order with which the three changes occurred. This study shows that gene inactivation or loss can be involved in the origin of phenotypic differences between species, thereby restricting the possibility of reversion to the ancestral state. It also demonstrates that parallel evolution of red flowers in three different species occurs via a common developmental/regulatory change but by mutations in different genes.
Figures
References
-
- Bull JJ, Charnov EL. On irreversible evolution. Evolution. 1985;39:1149–1155. - PubMed
-
- Cai DG, Kleine M, Kifle S, et al. (13 co-authors) Positional cloning of a gene for nematode resistance in sugar beet. Science. 1997;275:832–834. - PubMed
-
- Chen S, Matsubara K, Kokubun H, Kodama H, Watanabe H, Marchesi E, Ando T. Reconstructing historical events that occurred in the petunia Hf1 gene, which governs anthocyanin biosynthesis, and effects of artificial selection by breeding. Breed Sci. 2007;57:203–211.
-
- Colosimo PF, Hosemann KE, Balabhadra S, Villarreal G, Dickson M, Grimwood J, Schmutz J, Myers RM, Schluter D, Kingsley DM. Widespread parallel evolution in sticklebacks by repeated fixation of ectodysplasin alleles. Science. 2005;307:1928–1933. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
