

Clathrin mediates endocytosis and polar distribution of PIN auxin transporters in Arabidopsis
- PMID: 21551390
- PMCID: PMC3123958
- DOI: 10.1105/tpc.111.083030
Clathrin mediates endocytosis and polar distribution of PIN auxin transporters in Arabidopsis
Abstract
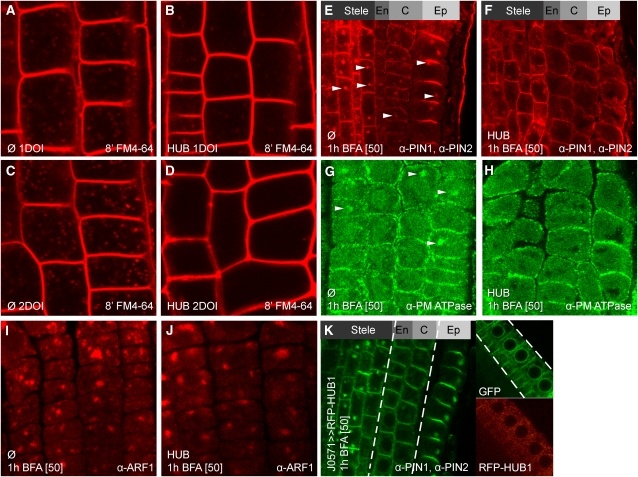
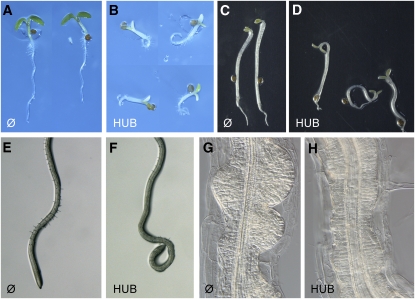
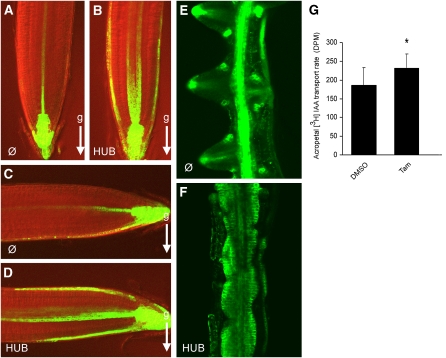
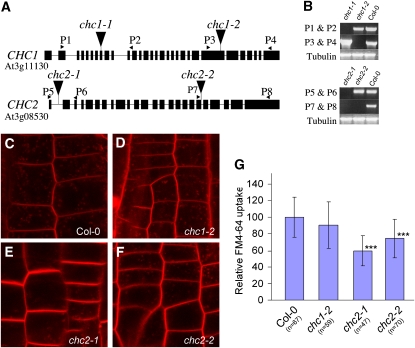
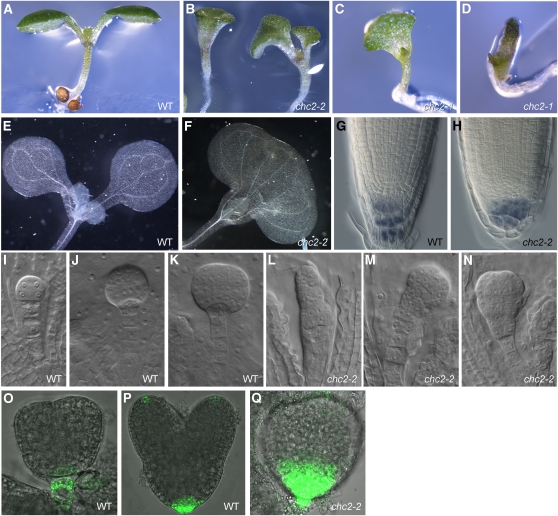
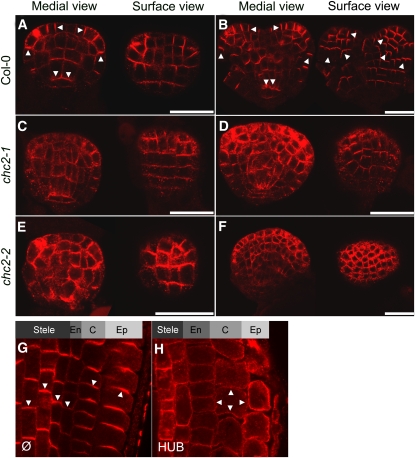
Endocytosis is a crucial mechanism by which eukaryotic cells internalize extracellular and plasma membrane material, and it is required for a multitude of cellular and developmental processes in unicellular and multicellular organisms. In animals and yeast, the best characterized pathway for endocytosis depends on the function of the vesicle coat protein clathrin. Clathrin-mediated endocytosis has recently been demonstrated also in plant cells, but its physiological and developmental roles remain unclear. Here, we assessed the roles of the clathrin-mediated mechanism of endocytosis in plants by genetic means. We interfered with clathrin heavy chain (CHC) function through mutants and dominant-negative approaches in Arabidopsis thaliana and established tools to manipulate clathrin function in a cell type-specific manner. The chc2 single mutants and dominant-negative CHC1 (HUB) transgenic lines were defective in bulk endocytosis as well as in internalization of prominent plasma membrane proteins. Interference with clathrin-mediated endocytosis led to defects in constitutive endocytic recycling of PIN auxin transporters and their polar distribution in embryos and roots. Consistent with this, these lines had altered auxin distribution patterns and associated auxin transport-related phenotypes, such as aberrant embryo patterning, imperfect cotyledon specification, agravitropic growth, and impaired lateral root organogenesis. Together, these data demonstrate a fundamental role for clathrin function in cell polarity, growth, patterning, and organogenesis in plants.
Figures
Similar articles
-
Clathrin-mediated constitutive endocytosis of PIN auxin efflux carriers in Arabidopsis.Curr Biol. 2007 Mar 20;17(6):520-7. doi: 10.1016/j.cub.2007.01.052. Epub 2007 Feb 15. Curr Biol. 2007. PMID: 17306539
-
Recycling, clustering, and endocytosis jointly maintain PIN auxin carrier polarity at the plasma membrane.Mol Syst Biol. 2011 Oct 25;7:540. doi: 10.1038/msb.2011.72. Mol Syst Biol. 2011. PMID: 22027551 Free PMC article.
-
ABP1 and ROP6 GTPase signaling regulate clathrin-mediated endocytosis in Arabidopsis roots.Curr Biol. 2012 Jul 24;22(14):1326-32. doi: 10.1016/j.cub.2012.05.020. Epub 2012 Jun 7. Curr Biol. 2012. PMID: 22683261
-
Budding and braking news about clathrin-mediated endocytosis.Curr Opin Plant Biol. 2013 Dec;16(6):718-25. doi: 10.1016/j.pbi.2013.09.005. Epub 2013 Oct 15. Curr Opin Plant Biol. 2013. PMID: 24139529 Review.
-
Polar targeting and endocytic recycling in auxin-dependent plant development.Annu Rev Cell Dev Biol. 2008;24:447-73. doi: 10.1146/annurev.cellbio.24.110707.175254. Annu Rev Cell Dev Biol. 2008. PMID: 18837671 Review.
Cited by
-
The sorting of cargo proteins in the plant trans-Golgi network.Front Plant Sci. 2022 Aug 11;13:957995. doi: 10.3389/fpls.2022.957995. eCollection 2022. Front Plant Sci. 2022. PMID: 36035717 Free PMC article. Review.
-
Clathrin-dependent endocytosis is required for immunity mediated by pattern recognition receptor kinases.Proc Natl Acad Sci U S A. 2016 Sep 27;113(39):11034-9. doi: 10.1073/pnas.1606004113. Epub 2016 Sep 20. Proc Natl Acad Sci U S A. 2016. PMID: 27651493 Free PMC article.
-
Danger-associated peptide signaling in Arabidopsis requires clathrin.Proc Natl Acad Sci U S A. 2016 Sep 27;113(39):11028-33. doi: 10.1073/pnas.1605588113. Epub 2016 Sep 20. Proc Natl Acad Sci U S A. 2016. PMID: 27651494 Free PMC article.
-
Evolutionary analysis of the ENTH/ANTH/VHS protein superfamily reveals a coevolution between membrane trafficking and metabolism.BMC Genomics. 2012 Jul 2;13:297. doi: 10.1186/1471-2164-13-297. BMC Genomics. 2012. PMID: 22748146 Free PMC article.
-
Salt-induced remodeling of spatially restricted clathrin-independent endocytic pathways in Arabidopsis root.Plant Cell. 2015 Apr;27(4):1297-315. doi: 10.1105/tpc.15.00154. Epub 2015 Apr 21. Plant Cell. 2015. PMID: 25901088 Free PMC article.
References
-
- Abas L., Benjamins R., Malenica N., Paciorek T., Wiśniewska J., Moulinier-Anzola J.C., Sieberer T., Friml J., Luschnig C. (2006). Intracellular trafficking and proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved in root gravitropism. Nat. Cell Biol. 8: 249–256 Erratum. Nat. Cell Biol. 8: 424 - PubMed
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Benjamins R., Quint A., Weijers D., Hooykaas P.J., Offringa R. (2001). The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport. Development 128: 4057–4067 - PubMed
-
- Benková E., Michniewicz M., Sauer M., Teichmann T., Seifertová D., Jürgens G., Friml J. (2003). Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115: 591–602 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
