

Nemo kinase phosphorylates β-catenin to promote ommatidial rotation and connects core PCP factors to E-cadherin-β-catenin
- PMID: 21552260
- PMCID: PMC3109122
- DOI: 10.1038/nsmb.2049
Nemo kinase phosphorylates β-catenin to promote ommatidial rotation and connects core PCP factors to E-cadherin-β-catenin
Abstract
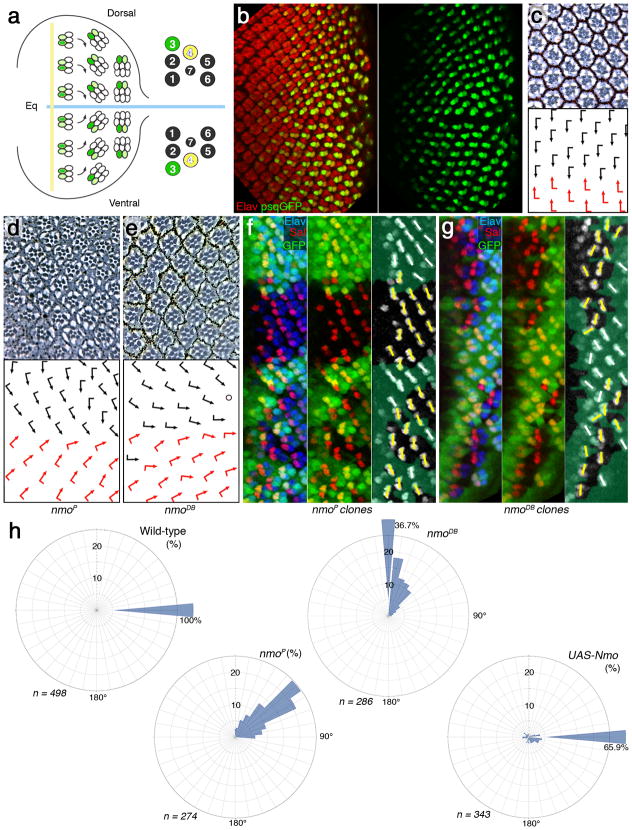
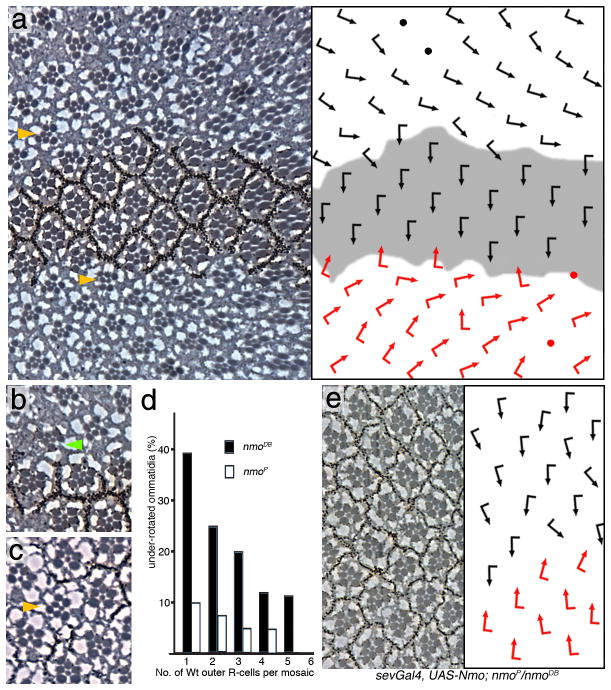
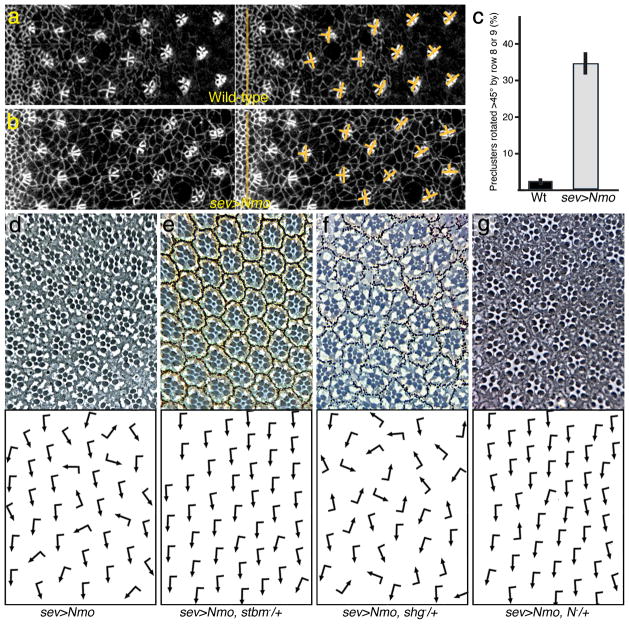
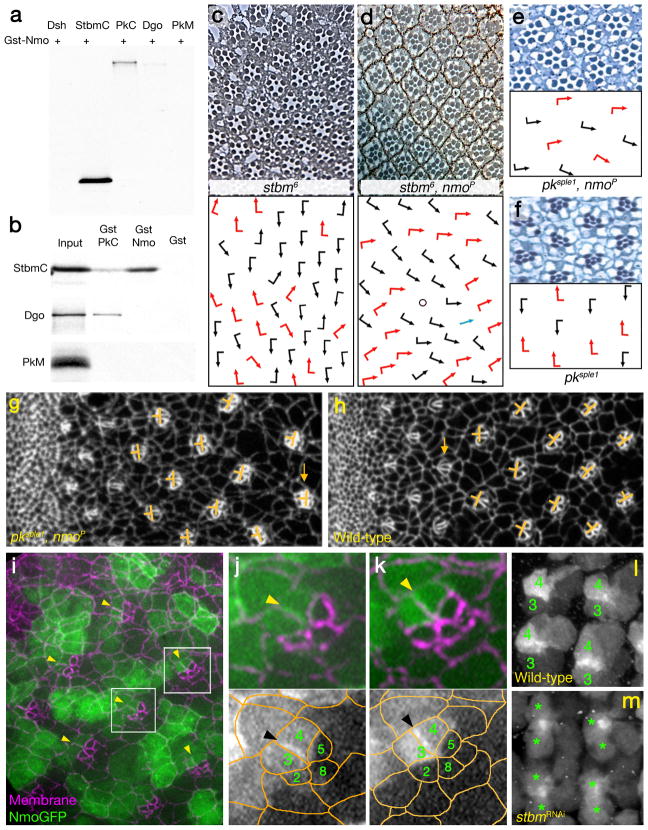
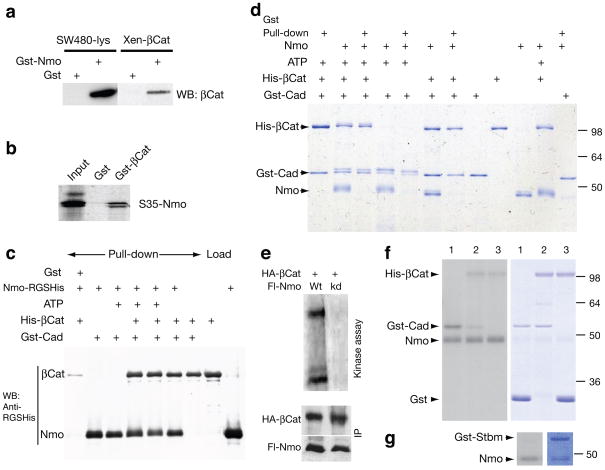
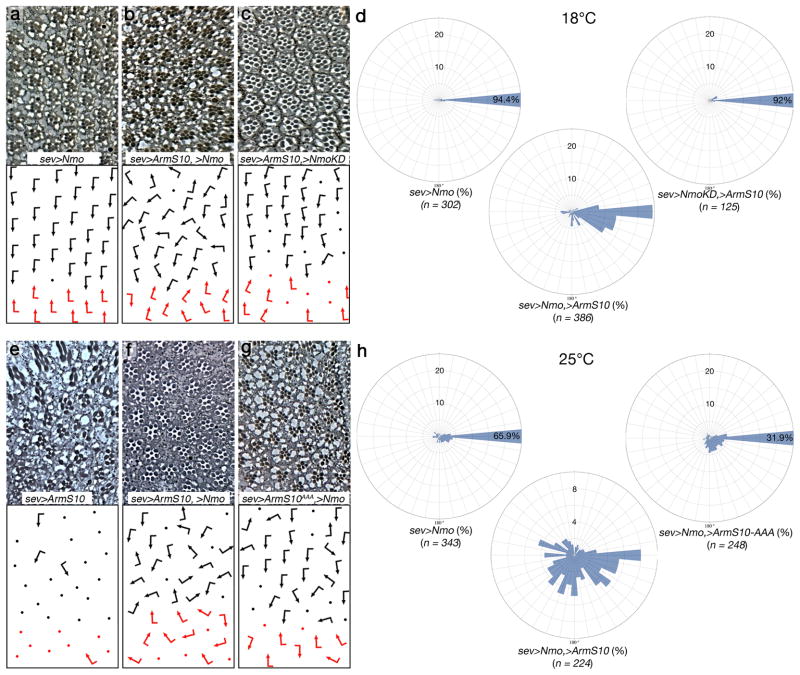
Frizzled planar cell polarity (PCP) signaling regulates cell motility in several tissues, including ommatidial rotation in Drosophila melanogaster. The Nemo kinase (Nlk in vertebrates) has also been linked to cell-motility regulation and ommatidial rotation but its mechanistic role(s) during rotation remain obscure. We show that nemo functions throughout the entire rotation movement, increasing the rotation rate. Genetic and molecular studies indicate that Nemo binds both the core PCP factor complex of Strabismus-Prickle, as well as the E-cadherin-β-catenin (E-cadherin-Armadillo in Drosophila) complex. These two complexes colocalize and, like Nemo, also promote rotation. Strabismus (also called Vang) binds and stabilizes Nemo asymmetrically within the ommatidial precluster; Nemo and β-catenin then act synergistically to promote rotation, which is mediated in vivo by Nemo's phosphorylation of β-catenin. Our data suggest that Nemo serves as a conserved molecular link between core PCP factors and E-cadherin-β-catenin complexes, promoting cell motility.
Figures
References
-
- Wolff T, Ready DF. Pattern formation in the Drosophila retina. In: Martinez-Arias MBA, editor. The development of Drosophila melanogaster. Cold Spring Harbor Press; Cold Spring Harbor: 1993. pp. 1277–1326.
-
- Seifert JR, Mlodzik M. Frizzled/PCP signalling: a conserved mechanism regulating cell polarity and directed motility. Nat Rev Genet. 2007;8:126–38. - PubMed
-
- Wang Y, Nathans J. Tissue/planar cell polarity in vertebrates: new insights and new questions. Development. 2007;134:647–58. - PubMed
-
- Mirkovic I, Mlodzik M. Cooperative activities of drosophila DE-cadherin and DN-cadherin regulate the cell motility process of ommatidial rotation. Development. 2006;133:3283–93. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
