

Multimeric assembly and biochemical characterization of the Trax-translin endonuclease complex
- PMID: 21552261
- PMCID: PMC3109869
- DOI: 10.1038/nsmb.2069
Multimeric assembly and biochemical characterization of the Trax-translin endonuclease complex
Abstract
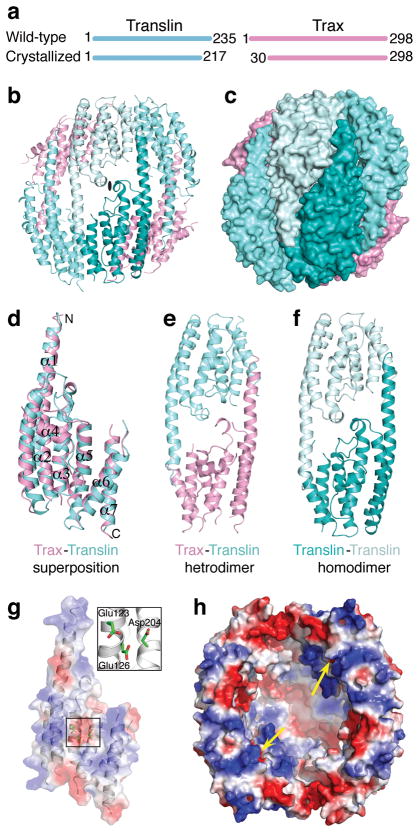
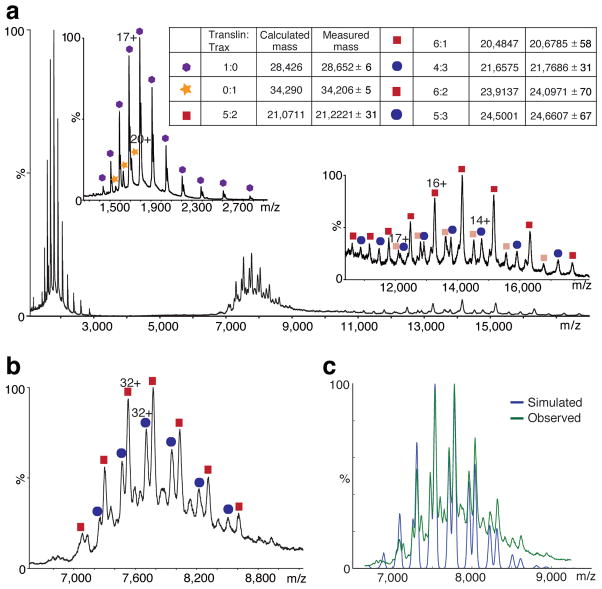
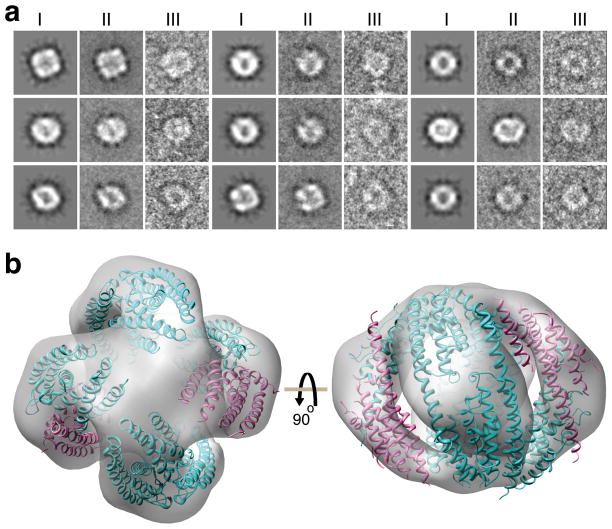
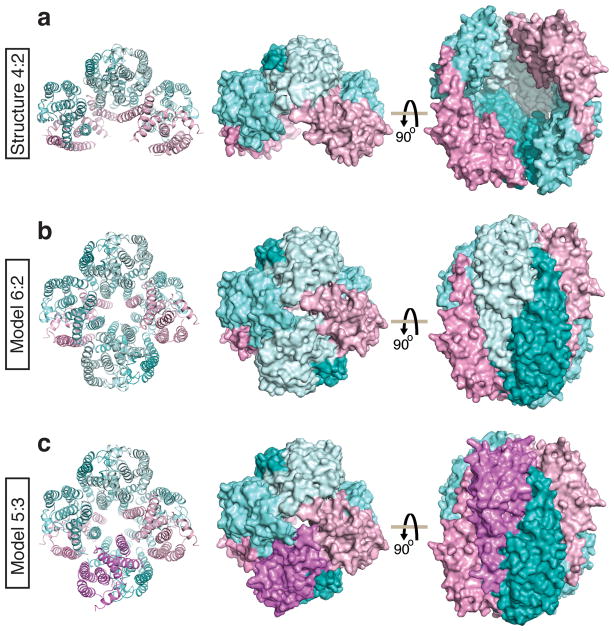
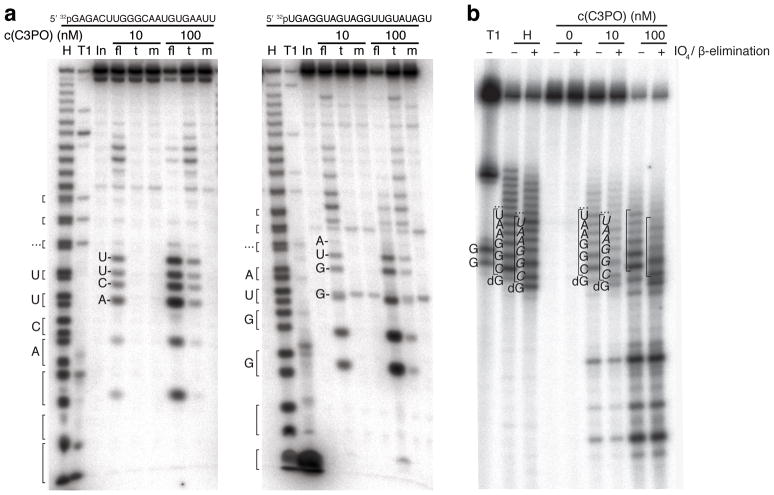
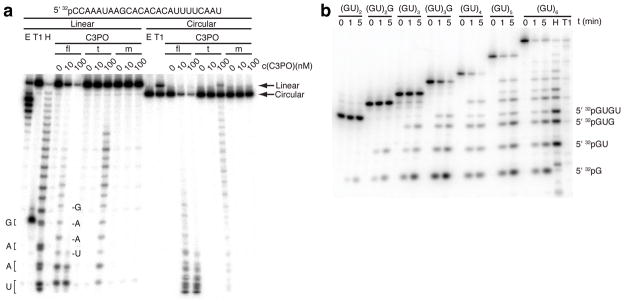
Trax-translin heteromers, also known as C3PO, have been proposed to activate the RNA-induced silencing complex (RISC) by facilitating endonucleolytic cleavage of the siRNA passenger strand. We report on the crystal structure of hexameric Drosophila C3PO formed by truncated translin and Trax, along with electron microscopic and mass spectrometric studies on octameric C3PO formed by full-length translin and Trax. Our studies establish that Trax adopts the translin fold, possesses catalytic centers essential for C3PO's endoRNase activity and interacts extensively with translin to form an octameric assembly. The catalytic pockets of Trax subunits are located within the interior chamber of the octameric scaffold. Truncated C3PO, like full-length C3PO, shows endoRNase activity that leaves 3'-hydroxyl-cleaved ends. We have measured the catalytic activity of C3PO and shown it to cleave almost stoichiometric amounts of substrate per second.
Figures
References
-
- Kim VN, Han J, Siomi MC. Biogenesis of small RNAs in animals. Nat Rev Mol Cell Biol. 2009;10:126–39. - PubMed
-
- Liu Q, et al. R2D2, a bridge between the initiation and effector steps of the Drosophila RNAi pathway. Science. 2003;301:1921–5. - PubMed
-
- Tomari Y, Matranga C, Haley B, Martinez N, Zamore PD. A protein sensor for siRNA asymmetry. Science. 2004;306:1377–80. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
