

Dysregulated TRK signalling is a therapeutic target in CYLD defective tumours
- PMID: 21552290
- PMCID: PMC3175103
- DOI: 10.1038/onc.2011.133
Dysregulated TRK signalling is a therapeutic target in CYLD defective tumours
Abstract
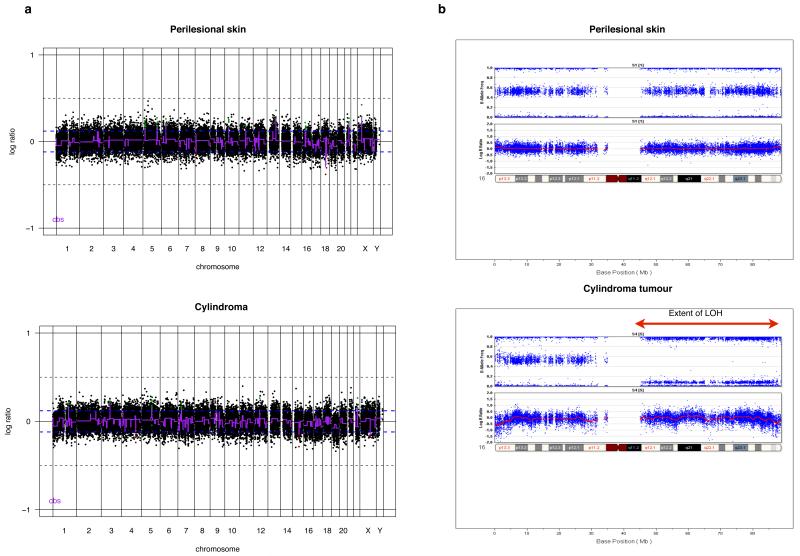
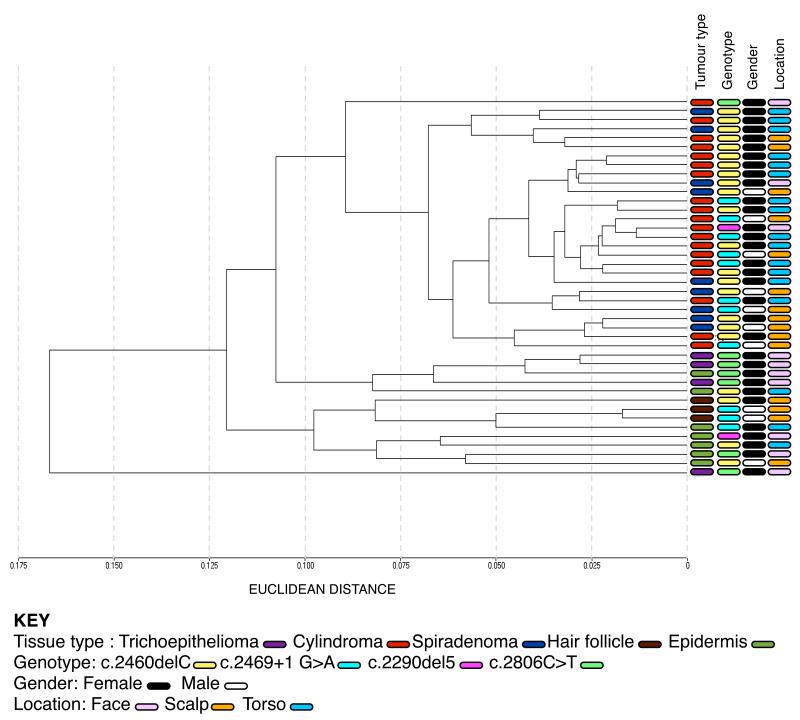
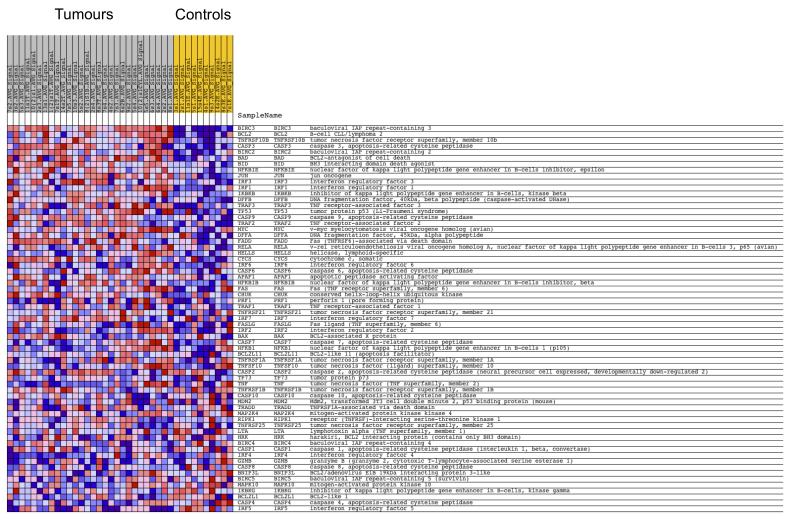
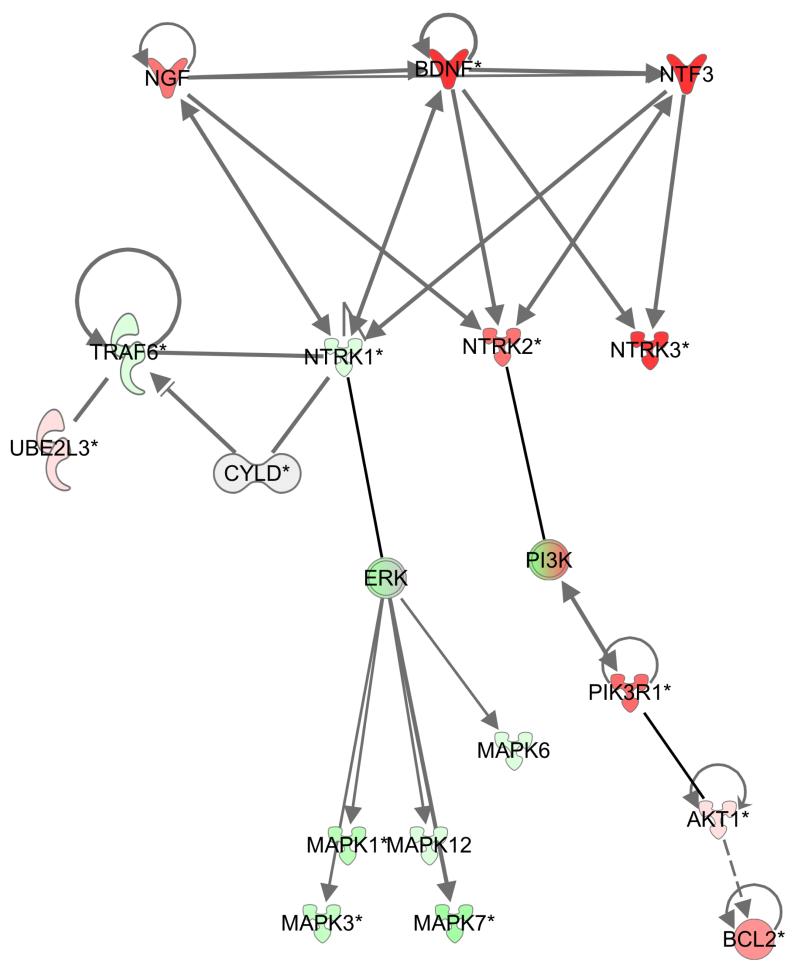
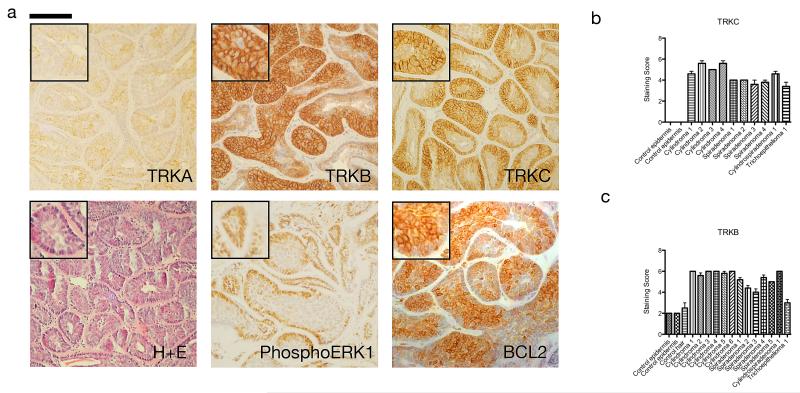
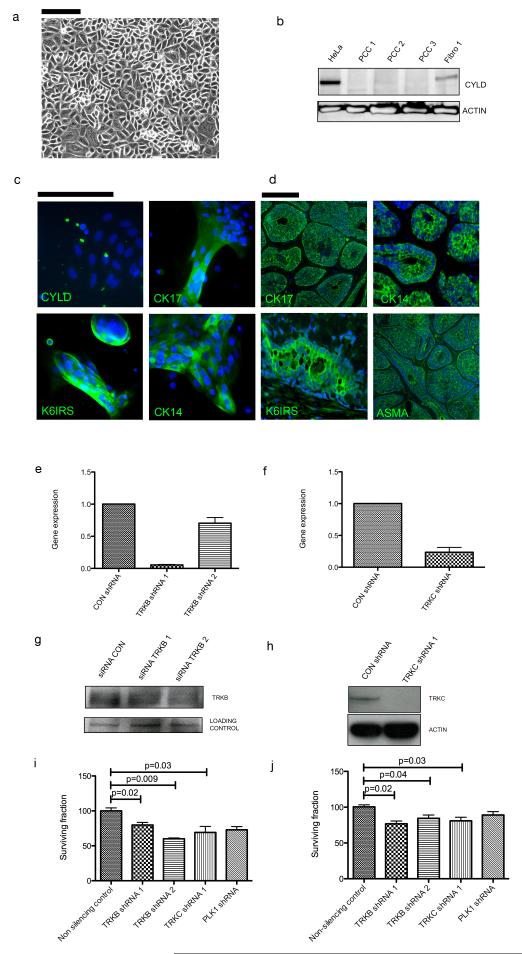
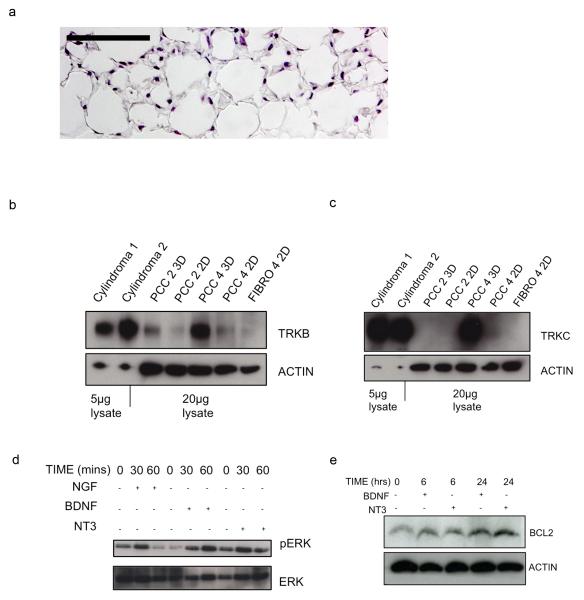
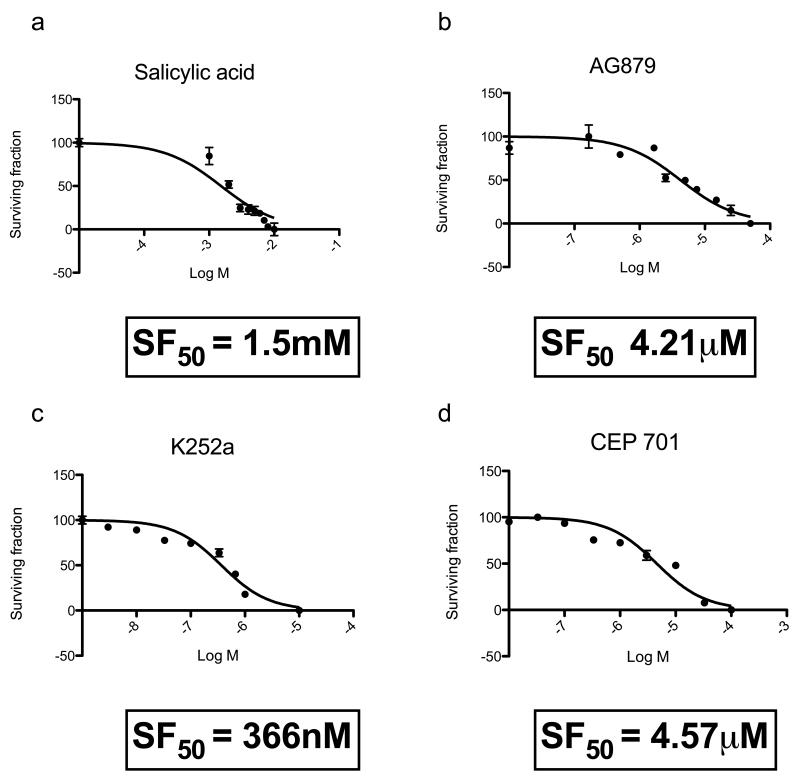
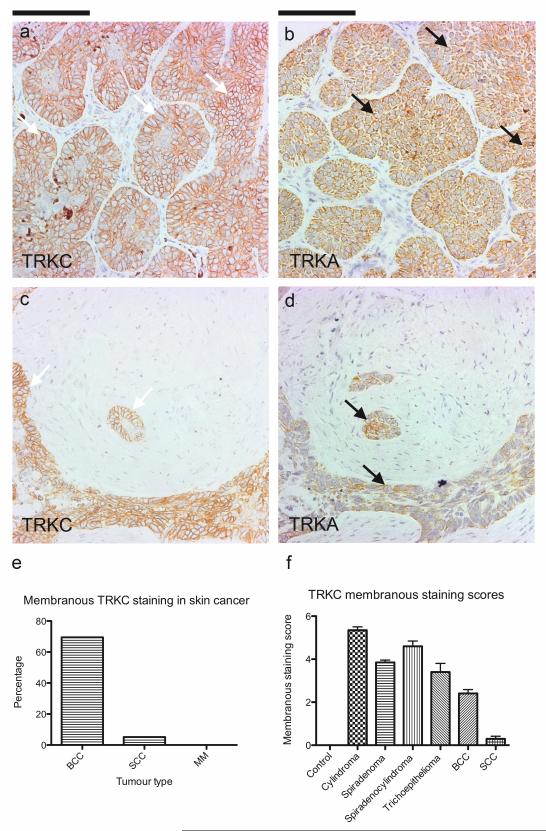
Individuals with germline mutations in the tumour-suppressor gene CYLD are at high risk of developing disfiguring cutaneous appendageal tumours, the defining tumour being the highly organised cylindroma. Here, we analysed CYLD mutant tumour genomes by array comparative genomic hybridisation and gene expression microarray analysis. CYLD mutant tumours were characterised by an absence of copy-number aberrations apart from LOH chromosome 16q, the genomic location of the CYLD gene. Gene expression profiling of CYLD mutant tumours showed dysregulated tropomyosin kinase (TRK) signalling, with overexpression of TRKB and TRKC in tumours when compared with perilesional skin. Immunohistochemical analysis of a tumour microarray showed strong membranous TRKB and TRKC staining in cylindromas, as well as elevated levels of ERK phosphorylation and BCL2 expression. Membranous TRKC overexpression was also observed in 70% of sporadic BCCs. RNA interference-mediated silencing of TRKB and TRKC, as well as treatment with the small-molecule TRK inhibitor lestaurtinib, reduced colony formation and proliferation in 3D primary cell cultures established from CYLD mutant tumours. These results suggest that TRK inhibition could be used as a strategy to treat tumours with loss of functional CYLD.
Figures
References
-
- Adly MA, Assaf HA, Nada EA, Soliman M, Hussein M. Human scalp skin and hair follicles express neurotrophin-3 and its high-affinity receptor tyrosine kinase C, and show hair cycle-dependent alterations in expression. Br J Dermatol. 2005;153:514–20. - PubMed
-
- Alvarez-Rodríguez R, Barzi M, Berenguer J, Pons S. Bone morphogenetic protein 2 opposes Shh-mediated proliferation in cerebellar granule cells through a TIEG-1-based regulation of Nmyc. J Biol Chem. 2007;282:37170–80. - PubMed
-
- Arévalo JC, Waite J, Rajagopal R, Beyna M, Chen Z-Y, Lee FS, et al. Cell survival through Trk neurotrophin receptors is differentially regulated by ubiquitination. Neuron. 2006;50:549–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
