

Review
doi: 10.1186/gb-2011-12-4-221.
Epub 2011 Apr 28.
Vive la différence: biogenesis and evolution of microRNAs in plants and animals
Affiliations
- PMID: 21554756
- PMCID: PMC3218855
- DOI: 10.1186/gb-2011-12-4-221
Item in Clipboard
Review
Vive la différence: biogenesis and evolution of microRNAs in plants and animals
Genome Biol.
2011.
Abstract
MicroRNAs are pervasive in both plants and animals, but many aspects of their biogenesis, function and evolution differ. We reveal how these differences contribute to characteristic features of microRNA evolution in the two kingdoms.
© 2011 BioMed Central Ltd
Figures
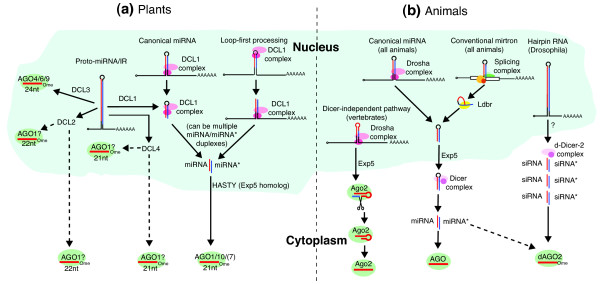
Major biogenesis pathways of small RNAs from inverted repeat transcripts in plants and animals. (a) In plants, canonical microRNAs (miRNAs) are produced by the nuclear RNase III Dicer-like1 (DCL1), which cuts from the base of the hairpin towards the loop; a subset of plant miRNAs are processed from the loop towards the hairpin. One miRNA/miRNA* duplex is shown, but there can be several such duplexes depending on the length of the stem. These are transported from the nucleus via HASTY, an Exportin5 (Exp5) homolog, for loading into an Argonaute (AGO) complex. The main miRNA effector in plants is AGO1, and to a lesser extent AGO10 and other AGOs; AGO7 carries the exceptional miRNA miR390. Long well-paired hairpins (proto-miR/inverted repeat (IR) transcripts) can be processed by a diversity of Dicers to generate either miRNAs or small interfering RNAs (siRNAs). The subcellular location for dicing by DCL2 and DCL4, and subsequent AGO loading of the resulting siRNAs, is not yet clear. nt, nucleotide. (b) In animals, canonical miRNAs are processed by the nuclear RNase III enzyme Drosha. The precursor miRNA (pre-miRNA) hairpin is exported to the cytoplasm by Exp5 to generate a single miRNA/miRNA* duplex, which is loaded into a miRNA class AGO protein (Drosophila dAGO1, Caenorhabditis elegans ALG1/2, or vertebrate Ago1 to Ago4). There are Drosha-independent non-canonical pathways, including the mirtron pathway where intron splicing and lariat debranching generate pre-miRNA hairpins. Also, vertebrate miR-451 is matured by a Dicer-independent route. Here, Drosha cleavage generates a short hairpin that is loaded into the 'Slicer' Ago2, which cleaves its 3' arm; this is resected to yield the mature miRNA. Unlike other vertebrate miRNAs, miR-451 can only be matured in Ago2. Finally, in the Drosophila hairpin RNA pathway, long inverted repeats are processed by the endogenous siRNA pathway, being cleaved by d-Dicer2 to generate siRNAs that load dAGO2. Many Drosophila miRNA* species are also preferentially sorted into dAGO2 (dashed arrow).
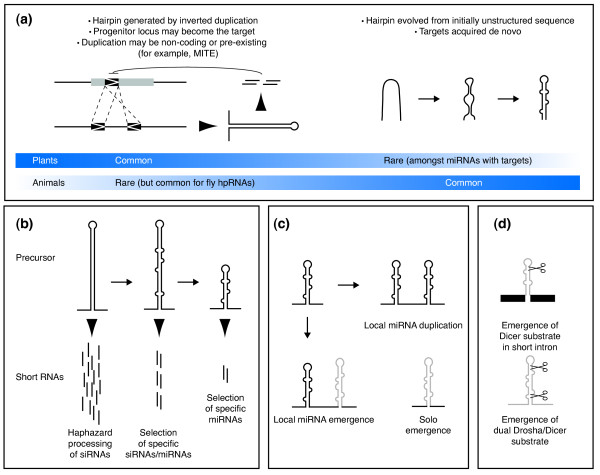
Modes of microRNA emergence in plants and animals. (a) Left: intragenomic duplications of protein-coding genes (or non-coding regions) can generate long foldbacks, which can be diced into small RNAs capable of targeting the progenitor transcript. This phenomenon seems common in plants, where extensive target complementarity is the rule, and ancestral relationships between microRNAs (miRNAs) and their targets can sometimes be detected; Drosophila hairpin RNA (hpRNA) may emerge similarly. MITE, miniature inverted-repeat transposable element. Right: inverted repeats might also emerge from initially unstructured sequences. This appears to be the dominant mode of miRNA emergence in animals. It also occurs in plants, but only rarely do such miRNAs appear to acquire functional targets. (b) Inferred model for plant miRNA emergence from long foldbacks; arrows indicate evolutionary relationships, arrowheads indicate small RNAs produced from a given hairpin. Long hairpins are processed haphazardly, often by different Dicers, to generate heterogeneous small interfering RNAs (siRNAs). As regulatory relationships are refined, the precision and phasing of hairpin processing may increase. Shortening of the hairpin to produce a single defined duplex may represent a mature state of plant miRNA evolution. (c) Expansion of miRNA clusters. In both plants and animals, local duplication may increase the dosage of a given miRNA. In animals, there may be an advantage for Drosha cleavage of hairpins emerging near extant miRNAs, leading to operons of unrelated miRNAs. (d) Different biogenesis mechanisms impose distinct demands on gene birth. Mirtrons need only evolve the capacity for one RNase III cleavage by Dicer, whereas canonical miRNAs need to gain the ability to be cleaved consecutively by Drosha and Dicer.
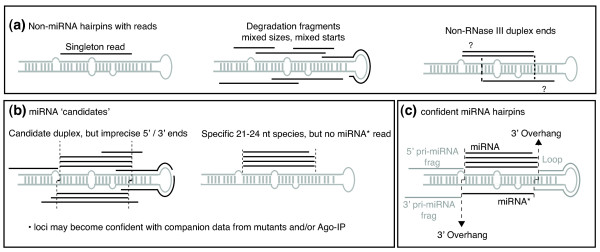
The complexity of annotating microRNAs from reads mapped to predicted hairpins. (a) Examples of loci that should not be annotated as microRNAs (miRNAs): hairpins with single reads, heterogeneously sized reads, and/or putatively duplexed reads lacking 3' overhangs. (b) Examples of loci with some, but insufficient, evidence for miRNA biogenesis; such loci are worth segregating as candidates pending further study. For instance, depletion of reads from miRNA biogenesis mutants or enrichment in Ago complexes could elevate their status from 'candidate' to 'confident'. It is also worth considering that candidate miRNA hairpins with relatively imprecise processing patterns may represent transitional intermediates in miRNA birth. nt, nucleotide. (c) Confident miRNA hairpins generate relatively precise miRNA/miRNA* duplexes with 3' overhangs. As datasets grow, it is often possible to observe cloned terminal loops or 5'/3' fragments (frag) of the primary miRNA (pri-miRNA) base, whose phasing with miRNA/miRNA* termini provide stringent evidence for in vivo cleavage reactions. Note that the vertebrate locus mir-451 matures by direct hairpin cleavage by Ago2, and not via a miRNA/miRNA* intermediate; thus, the criteria outlined in this figure are not applicable to mir-451-class substrates.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
