

Febuxostat inhibition of endothelial-bound XO: implications for targeting vascular ROS production
- PMID: 21554948
- PMCID: PMC3130629
- DOI: 10.1016/j.freeradbiomed.2011.04.004
Febuxostat inhibition of endothelial-bound XO: implications for targeting vascular ROS production
Abstract
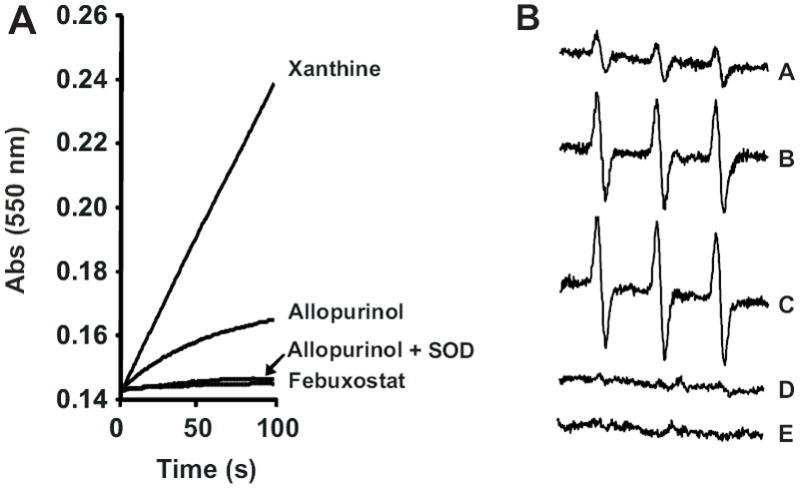
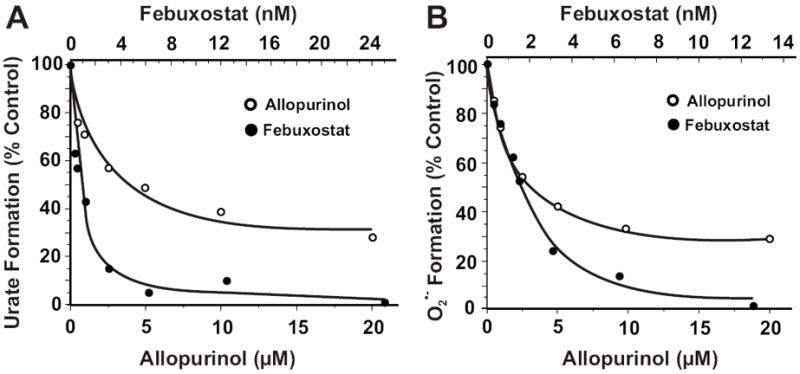
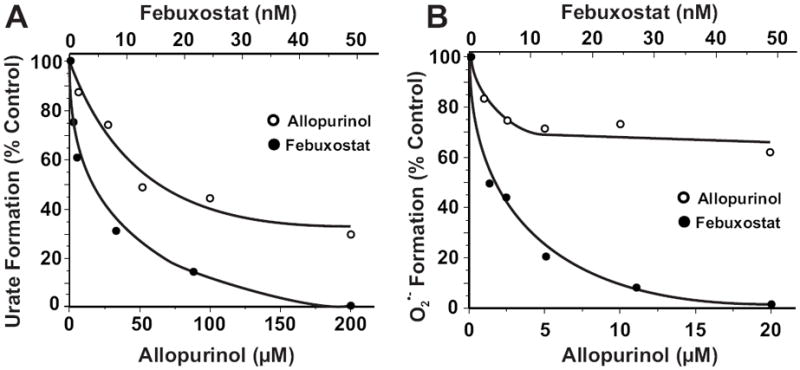
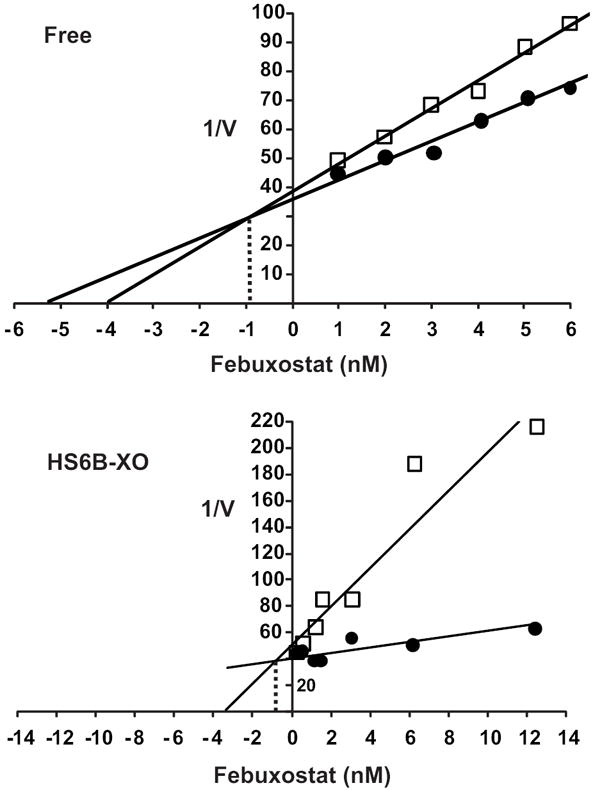
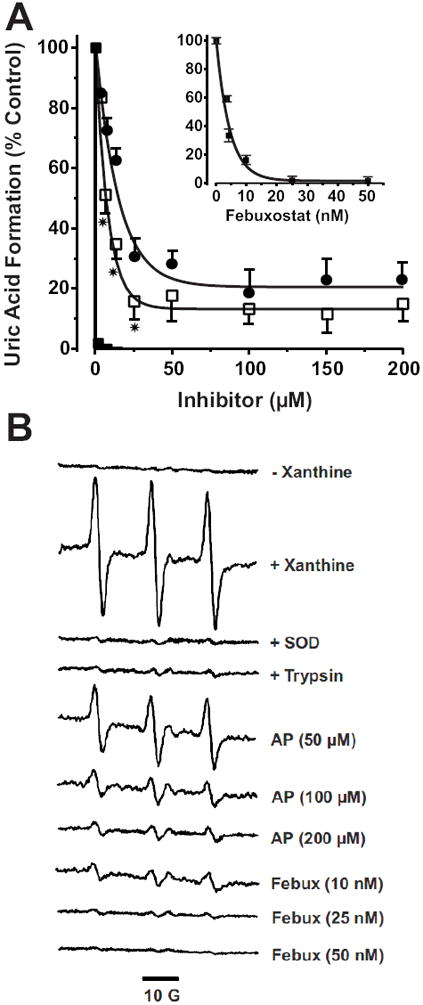
Xanthine oxidase (XO) is a critical source of reactive oxygen species (ROS) that contribute to vascular inflammation. Binding of XO to vascular endothelial cell glycosaminoglycans (GAGs) results in significant resistance to inhibition by traditional pyrazolopyrimidine-based inhibitors such as allopurinol. Therefore, we compared the extent of XO inhibition (free and GAG-bound) by allopurinol to that by febuxostat, a newly approved nonpurine XO-specific inhibitor. In solution, febuxostat was 1000-fold more potent than allopurinol at inhibiting XO-dependent uric acid formation (IC₅₀= 1.8 nM vs 2.9 μM). Association of XO with heparin-Sepharose 6B (HS6B-XO) had minimal effect on the inhibition of uric acid formation by febuxostat (IC₅₀= 4.4 nM) while further limiting the effect of allopurinol (IC₅₀= 64 μM). Kinetic analysis of febuxostat inhibition revealed K(i) values of 0.96 (free) and 0.92 nM (HS6B-XO), confirming equivalent inhibition for both free and GAG-immobilized enzyme. When XO was bound to endothelial cell GAGs, complete enzyme inhibition was observed with 25 nM febuxostat, whereas no more than 80% inhibition was seen with either allopurinol or oxypurinol, even at concentrations above those tolerated clinically. The superior potency for inhibition of endothelium-associated XO is predictive of a significant role for febuxostat in investigating pathological states in which XO-derived ROS are contributive and traditional XO inhibitors are only slightly effective.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Amaya Y, Yamazaki K, Sato M, Noda K, Nishino T. Proteolytic conversion of xanthine dehydrogenase from the NAD-dependent type to the O2-dependent type. Amino acid sequence of rat liver xanthine dehydrogenase and identification of the cleavage sites of the enzyme protein during irreversible conversion by trypsin. J Biol Chem. 1990;265:14170–14175. - PubMed
-
- Waud WR, Rajagopalan KV. The mechanism of conversion of rat liver xanthine dehydrogenase from an NAD+-dependent form (type D) to an O2-dependent form (type O) Arch Biochem Biophys. 1976;172:365–379. - PubMed
-
- Harris CM, Massey V. The reaction of reduced xanthine dehydrogenase with molecular oxygen. Reaction kinetics and measurment of superoxide radical. J Biol Chem. 1997;272:8370–8379. - PubMed
-
- Butler R, Morris AD, Belch JJ, Hill A, Struthers AD. Allopurinol normalizes endothelial dysfunction in type 2 diabetics with mild hypertension. Hypertension. 2000;35:746–751. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
