

Review
doi: 10.1016/j.neuron.2011.04.009.
The many faces of tau
Affiliations
- PMID: 21555069
- PMCID: PMC3319390
- DOI: 10.1016/j.neuron.2011.04.009
Item in Clipboard
Review
The many faces of tau
Neuron.
.
Abstract
While the microtubule-binding capacity of the protein tau has been known for many years, new functions of tau in signaling and cytoskeletal organization have recently emerged. In this review, we highlight these functions and the potential roles of tau in neurodegenerative disease. We also discuss the therapeutic potential of drugs targeting various aspects of tau biology.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
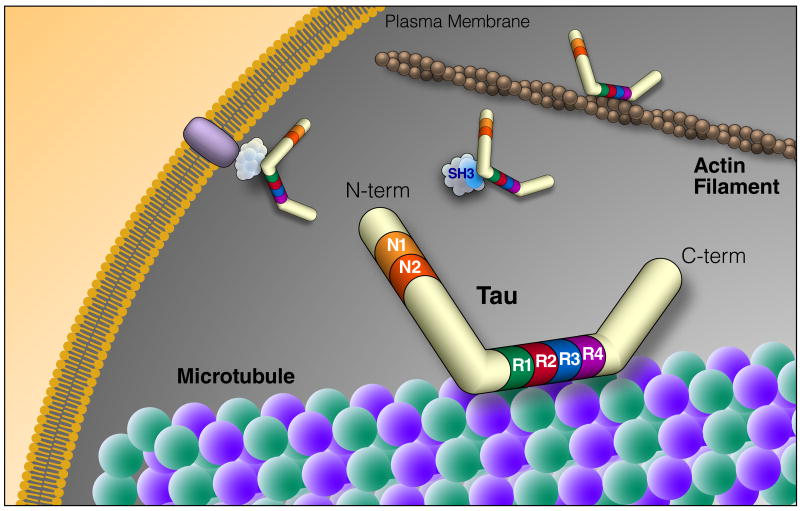
Tau is an intrinsically disordered protein that can be alternatively spliced at N terminal exons (N1, 2) and the microtubule repeat domains (R). The domains of tau bind many different types of molecules, suggesting a central role in signaling pathways and cytoskeletal organization. The diversity of tau binding partners is highlighted in Table 1. N-term, N-terminus; C-term, C-terminus; SH3, protein SH3 domain
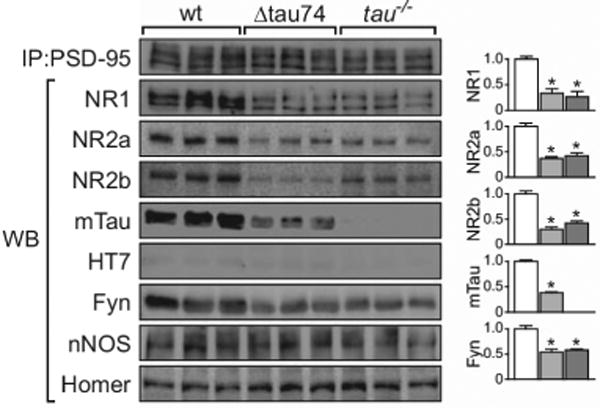
Tau immunoprecipitates with PSD95, a postsynaptic protein, and regulates the association of PSD-95 with NMDAR subunits and the tyrosine kinase Fyn. Overexpression of a human N-terminal truncation of tau (Δtau74, light grey) or knocking out tau (tau−/−, dark grey) reduced the amount of NR1, NR2a, NR2b, and Fyn within PSD95 complexes. From Ittner et al., 2010 with permission.
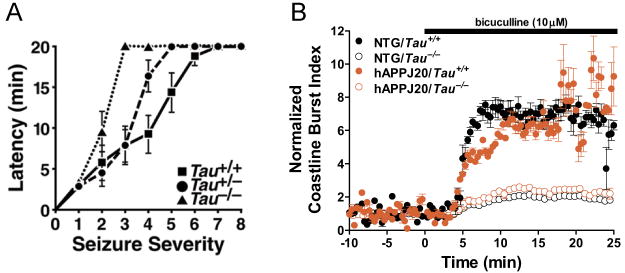
A) Partial or complete reduction of tau in mice without hAPP expression delayed the onset and reduced the severity of seizures induced by pentylenetetrazol, a GABAA receptor antagonist (Roberson et al., 2007). B) Tau knockout reduced aberrant neuronal discharges in acute hippocampal slices after disinhibition with the GABAA receptor antagonist bicuculline, as illustrated by measurements of the coastline burst index (Roberson et al., 2011). NTG, no hAPP expression; hAPPJ20, hAPP mice from line J20
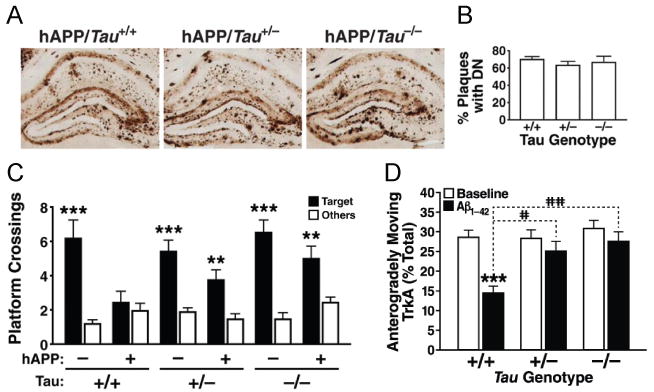
Tau reduction did not affect (A) Aβ deposition or (B) the number of plaques with dystrophic neurites (DN) in hAPP-J20 mice. However, even partial tau reduction prevented (C) memory deficits in the Morris water maze (72 hour probe trial) in hAPP-J20 mice (Roberson et al., 2007) and (D) Aβ oligomer-induced axonal transport deficits in primary hippocampal neurons, although it had no effect on axonal transport at baseline (Vossel et al., 2010).
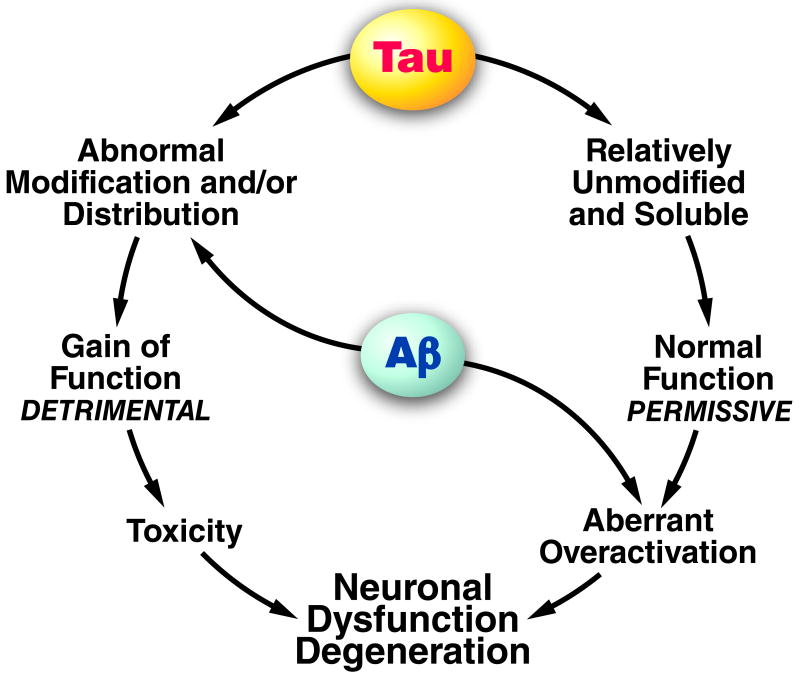
Experiments in which neurons were acutely exposed to Aβ suggest that Aβ can trigger tau-mediated neurotoxicity by enhancing tau phosphorylation, which in turn directs pathogenic tau species into dendritic spines where they exert adverse effects (left). In hAPP mice, though, in which neurons are chronically exposed to elevated Aβ levels, it has so far been impossible to find clear evidence for a similar process. Nonetheless, Aβ-induced neuronal dysfunction in these models strictly depends on the presence of tau, raising the possibility that physiological functions of tau permit Aβ to cause neuronal dysfunction (right). Such functions may involve the intraneuronal trafficking of factors that regulate synaptic activity at the pre- or postsynaptic level.
References
-
- Alonso AC, Grundke-Iqbal I, Iqbal K. Alzheimer's disease hyperphosphorylated tau sequesters normal tau into tangles of filaments and disassembles microtubules. Nat Med. 1996;2:783–787. - PubMed
-
- Alvarez A, Toro R, Cáceres A, Maccioni RB. Inhibition of tau phosphorylating protein kinase cdk5 prevents beta-amyloid-induced neuronal death. FEBS Lett. 1999;459:421–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
