

An antisense CAG repeat transcript at JPH3 locus mediates expanded polyglutamine protein toxicity in Huntington's disease-like 2 mice
- PMID: 21555070
- PMCID: PMC3107122
- DOI: 10.1016/j.neuron.2011.03.021
An antisense CAG repeat transcript at JPH3 locus mediates expanded polyglutamine protein toxicity in Huntington's disease-like 2 mice
Abstract
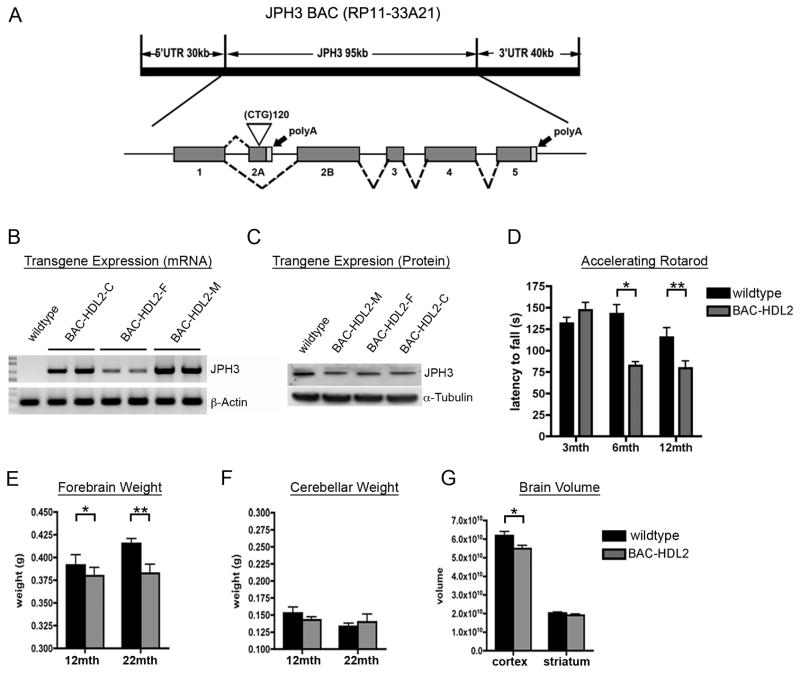
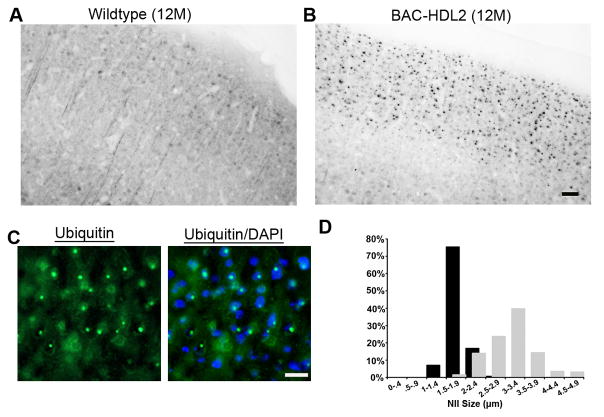
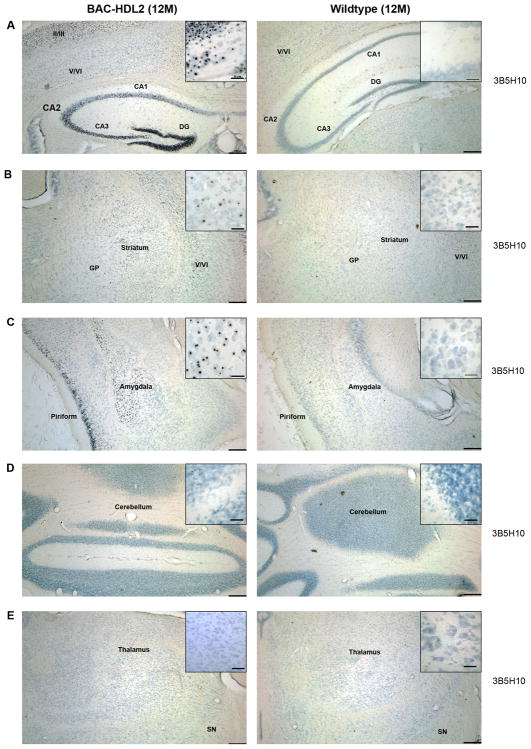
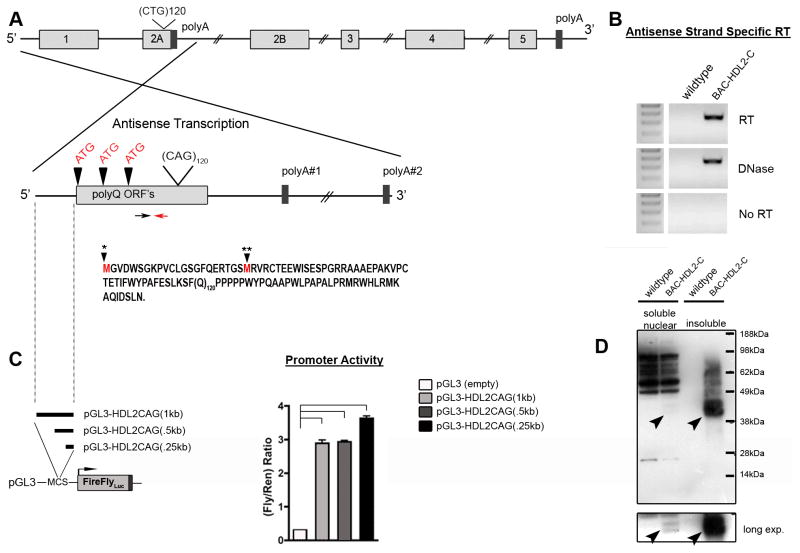
Huntington's disease-like-2 (HDL2) is a phenocopy of Huntington's disease caused by CTG/CAG repeat expansion at the Junctophilin-3 (JPH3) locus. The mechanisms underlying HDL2 pathogenesis remain unclear. Here we developed a BAC transgenic mouse model of HDL2 (BAC-HDL2) that exhibits progressive motor deficits, selective neurodegenerative pathology, and ubiquitin-positive nuclear inclusions (NIs). Molecular analyses reveal a promoter at the transgene locus driving the expression of a CAG repeat transcript (HDL2-CAG) from the strand antisense to JPH3, which encodes an expanded polyglutamine (polyQ) protein. Importantly, BAC-HDL2 mice, but not control BAC mice, accumulate polyQ-containing NIs in a pattern strikingly similar to those in the patients. Furthermore, BAC mice with genetic silencing of the expanded CUG transcript still express HDL2-CAG transcript and manifest polyQ pathogenesis. Finally, studies of HDL2 mice and patients revealed CBP sequestration into NIs and evidence for interference of CBP-mediated transcriptional activation. These results suggest overlapping polyQ-mediated pathogenic mechanisms in HD and HDL2.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
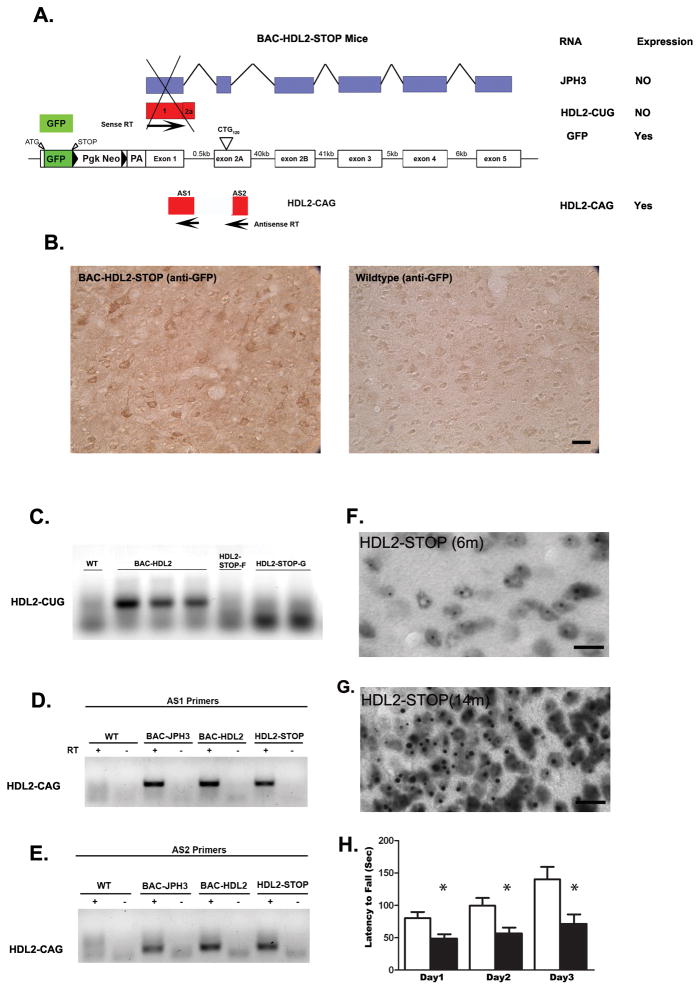
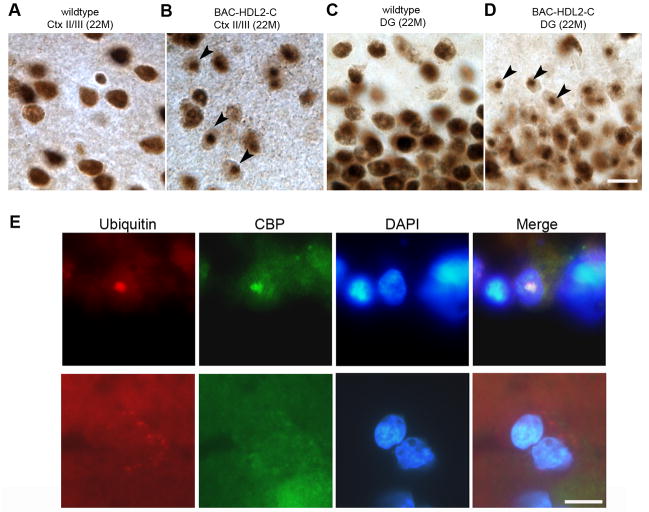
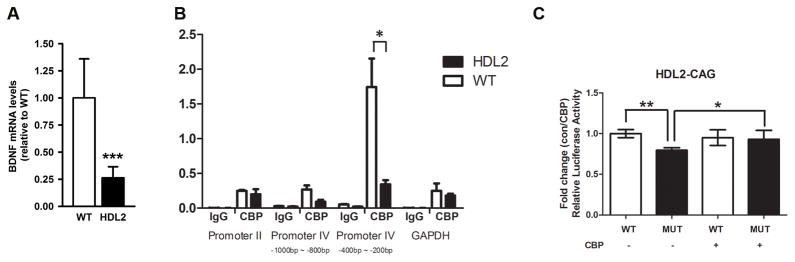
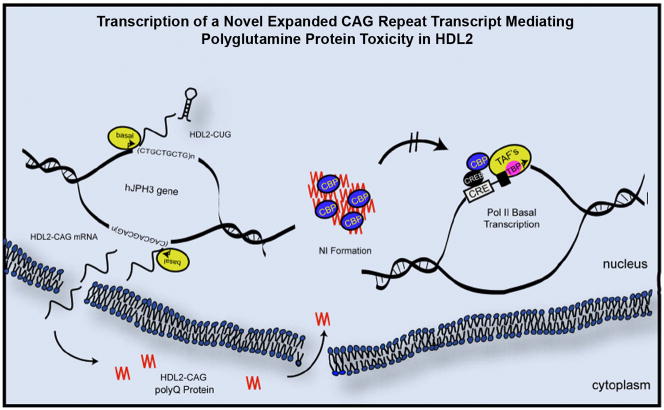
Comment in
-
Are polyglutamine diseases expanding?Neuron. 2011 May 12;70(3):377-8. doi: 10.1016/j.neuron.2011.04.015. Neuron. 2011. PMID: 21555065
References
-
- Brooks E, Arrasate M, Cheung K, Finkbeiner SM. Using antibodies to analyze polyglutamine stretches. Methods Mol Biol. 2004;277:103–128. - PubMed
-
- Chan HM, La Thangue NB. p300/CBP proteins: HATs for transcriptional bridges and scaffolds. J Cell Sci. 2001;114:2363–2373. - PubMed
-
- Dorsman JC, Pepers B, Langenberg D, Kerkdijk H, Ijszenga M, den Dunnen JT, Roos RA, van Ommen GJ. Strong aggregation and increased toxicity of polyleucine over polyglutamine stretches in mammalian cells. Hum Mol Genet. 2002;11:1487–1496. - PubMed
-
- Gambazzi L, Gokce O, Seredenina T, Katsyuba E, Runne H, Markram H, Giugliano M, Luthi-Carter R. Diminished activity-dependent brain-derived neurotrophic factor expression underlies cortical neuron microcircuit hypoconnectivity resulting from exposure to mutant huntingtin fragments. J Pharmacol Exp Ther. 2010;335:13–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
