

Mechanistic logic underlying the axonal transport of cytosolic proteins
- PMID: 21555071
- PMCID: PMC3096075
- DOI: 10.1016/j.neuron.2011.03.022
Mechanistic logic underlying the axonal transport of cytosolic proteins
Abstract
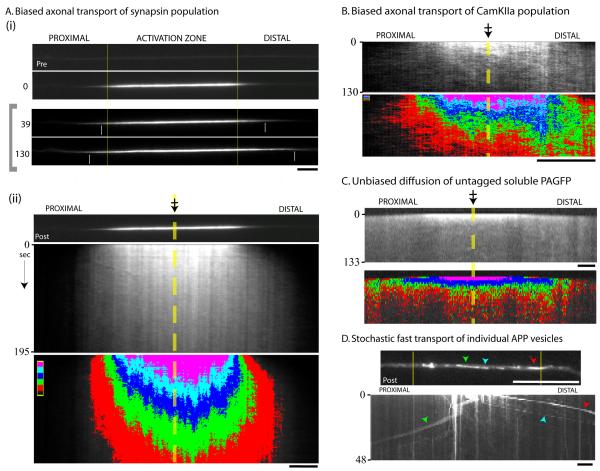
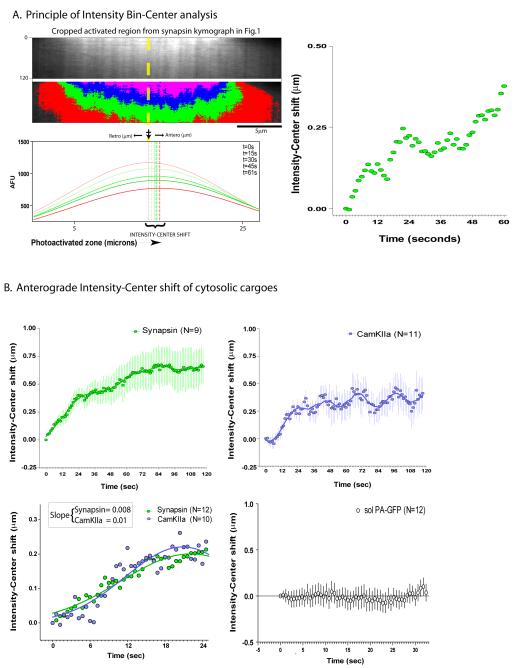
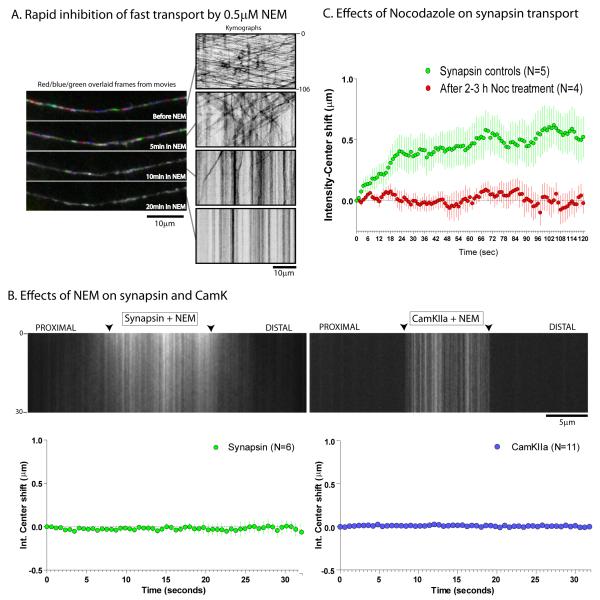
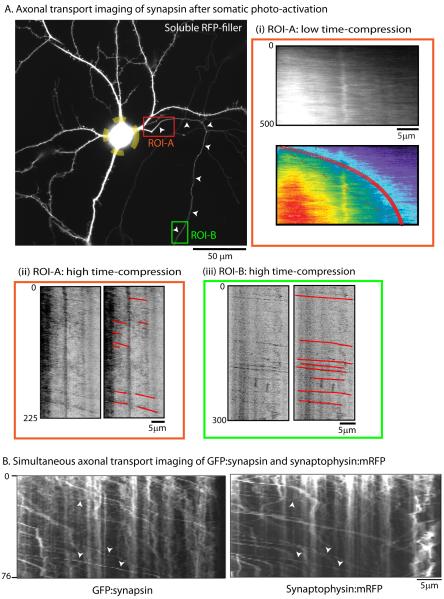
Proteins vital to presynaptic function are synthesized in the neuronal perikarya and delivered into synapses via two modes of axonal transport. While membrane-anchoring proteins are conveyed in fast axonal transport via motor-driven vesicles, cytosolic proteins travel in slow axonal transport via mechanisms that are poorly understood. We found that in cultured axons, populations of cytosolic proteins tagged to photoactivatable GFP (PAGFP) move with a slow motor-dependent anterograde bias distinct from both vesicular trafficking and diffusion of untagged PAGFP. The overall bias is likely generated by an intricate particle kinetics involving transient assembly and short-range vectorial spurts. In vivo biochemical studies reveal that cytosolic proteins are organized into higher order structures within axon-enriched fractions that are largely segregated from vesicles. Data-driven biophysical modeling best predicts a scenario where soluble molecules dynamically assemble into mobile supramolecular structures. We propose a model where cytosolic proteins are transported by dynamically assembling into multiprotein complexes that are directly/indirectly conveyed by motors.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
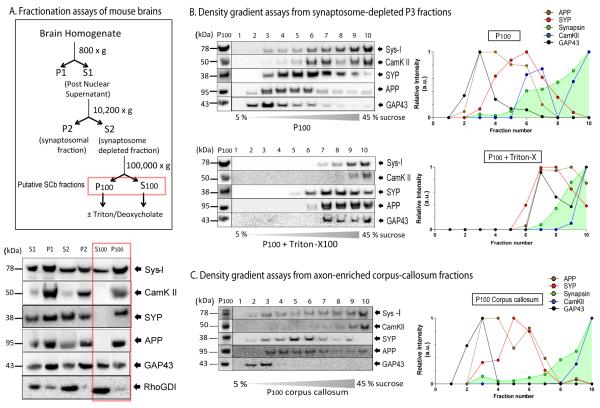
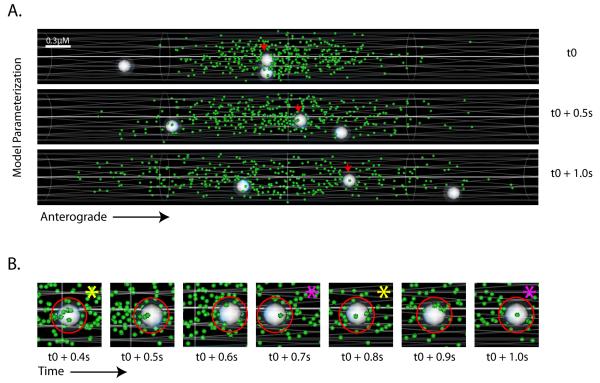
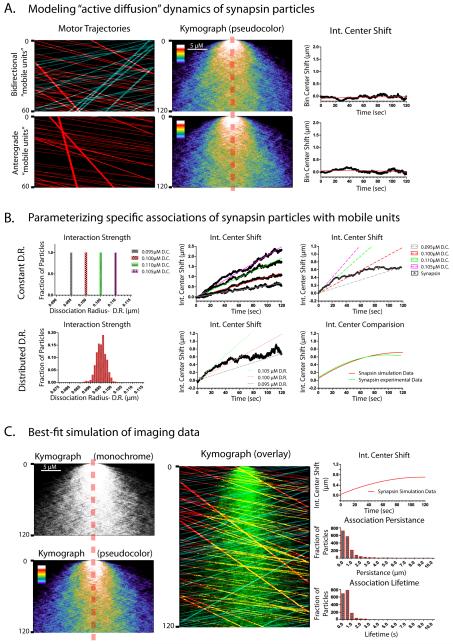
Comment in
-
The curious case of the soluble protein.Dev Cell. 2011 May 17;20(5):581-2. doi: 10.1016/j.devcel.2011.05.003. Dev Cell. 2011. PMID: 21571216
References
-
- Ahmari SE, Buchanan J, Smith SJ. Assembly of presynaptic active zones from cytoplasmic transport packets. Nat.Neurosci. 2000;3:445–451. - PubMed
-
- Booj S, Larsson PA, Dahllof AG, Dahlstrom A. Axonal transport of synapsin I- and cholinergic synaptic vesicle-like material; further immunohistochemical evidence for transport of axonal cholinergic transmitter vesicles in motor neurons. Acta Physiol Scand. 1986;128:155–165. - PubMed
-
- Brangwynne CP, Koenderink GH, MacKintosh FC, Weitz DA. Intracellular transport by active diffusion. Trends Cell Biol. 2009;19:423–427. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
