

Genome-wide detection of chromosomal rearrangements, indels, and mutations in circular chromosomes by short read sequencing
- PMID: 21555365
- PMCID: PMC3149504
- DOI: 10.1101/gr.117416.110
Genome-wide detection of chromosomal rearrangements, indels, and mutations in circular chromosomes by short read sequencing
Abstract
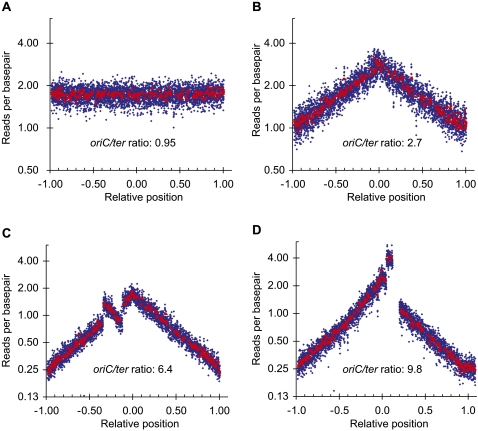
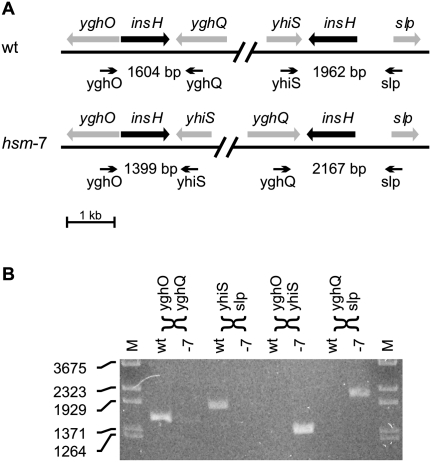
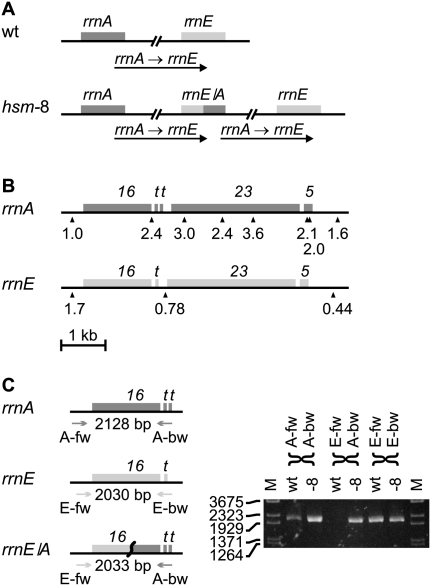
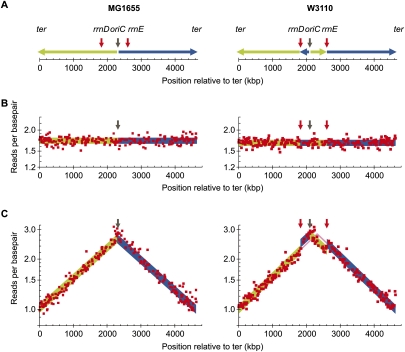
Whole-genome sequencing (WGS) with new short-read sequencing technologies has recently been applied for genome-wide identification of mutations. Genomic rearrangements have, however, often remained undetected by WGS, and additional analyses are required for their detection. Here, we have applied a combination of WGS and genome copy number analysis, for the identification of mutations that suppress the growth deficiency imposed by excessive initiations from the Escherichia coli origin of replication, oriC. The E. coli chromosome, like the majority of bacterial chromosomes, is circular, and DNA replication is initiated by assembling two replication complexes at the origin, oriC. These complexes then replicate the chromosome bidirectionally toward the terminus, ter. In a population of growing cells, this results in a copy number gradient, so that origin-proximal sequences are more frequent than origin-distal sequences. Major rearrangements in the chromosome are, therefore, readily identified by changes in copy number, i.e., certain sequences become over- or under-represented. Of the eight mutations analyzed in detail here, six were found to affect a single gene only, one was a large chromosomal inversion, and one was a large chromosomal duplication. The latter two mutations could not be detected solely by WGS, validating the present approach for identification of genomic rearrangements. We further suggest the use of copy number analysis in combination with WGS for validation of newly assembled bacterial chromosomes.
Figures
References
-
- Atlung T, Løbner-Olesen A, Hansen FG 1987. Overproduction of DnaA protein stimulates initiation of chromosome and minichromosome replication in E. coli. Mol Gen Genet 206: 51–59 - PubMed
-
- Barrick JE, Yu DS, Yoon SH, Jeong H, Oh TK, Schneider D, Lenski RE, Kim JF 2009. Genome evolution and adaptation in a long-term experiment with Escherichia coli. Nature 461: 1243–1247 - PubMed
-
- Blattner FR, Plunkett G III, Bloch CA, Perna NT, Burland V, Riley M, Collado-Vides J, Glasner JD, Rode CK, Mayhew GF, et al. 1997. The complete genome sequence of Escherichia coli K-12. Science 277: 1453–1462 - PubMed
-
- Bremer H, Churchward G 1977. An examination of the Cooper-Helmstetter theory of DNA replication in bacteria and its underlying assumptions. J Theor Biol 69: 645–654 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources