

Vibrio cholerae requires the type VI secretion system virulence factor VasX to kill Dictyostelium discoideum
- PMID: 21555399
- PMCID: PMC3191968
- DOI: 10.1128/IAI.01266-10
Vibrio cholerae requires the type VI secretion system virulence factor VasX to kill Dictyostelium discoideum
Abstract
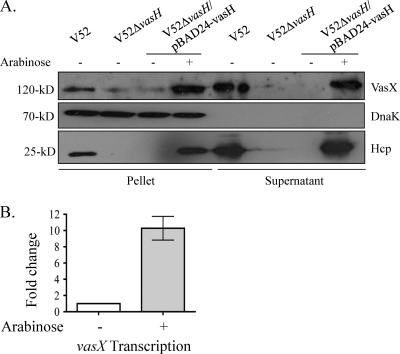
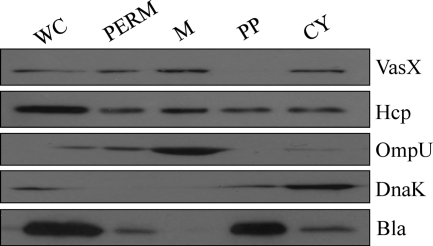
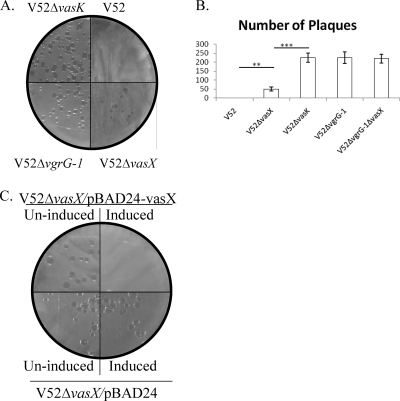
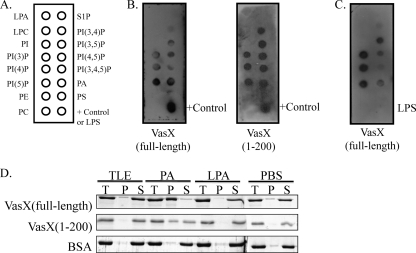
The type VI secretion system (T6SS) is recognized as an important virulence mechanism in several Gram-negative pathogens. In Vibrio cholerae, the causative agent of the diarrheal disease cholera, a minimum of three gene clusters--one main cluster and two auxiliary clusters--are required to form a functional T6SS apparatus capable of conferring virulence toward eukaryotic and prokaryotic hosts. Despite an increasing understanding of the components that make up the T6SS apparatus, little is known about the regulation of these genes and the gene products delivered by this nanomachine. VasH is an important regulator of the V. cholerae T6SS. Here, we present evidence that VasH regulates the production of a newly identified protein, VasX, which in turn requires a functional T6SS for secretion. Deletion of vasX does not affect export or enzymatic function of the structural T6SS proteins Hcp and VgrG-1, suggesting that VasX is dispensable for the assembly of the physical translocon complex. VasX localizes to the bacterial membrane and interacts with membrane lipids. We present VasX as a novel virulence factor of the T6SS, as a V. cholerae mutant lacking vasX exhibits a phenotype of attenuated virulence toward Dictyostelium discoideum.
Figures


References
-
- Arnold R. S., Cornell R. B. 1996. Lipid regulation of CTP: phosphocholine cytidylyltransferase: electrostatic, hydrophobic, and synergistic interactions of anionic phospholipids and diacylglycerol. Biochemistry 35:9917–9924 - PubMed
-
- Bennett-Lovsey R. M., Herbert A. D., Sternberg M. J., Kelley L. A. 2008. Exploring the extremes of sequence/structure space with ensemble fold recognition in the program Phyre. Proteins 70:611–625 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
