

Dual-mode of insulin action controls GLUT4 vesicle exocytosis
- PMID: 21555461
- PMCID: PMC3166865
- DOI: 10.1083/jcb.201008135
Dual-mode of insulin action controls GLUT4 vesicle exocytosis
Abstract
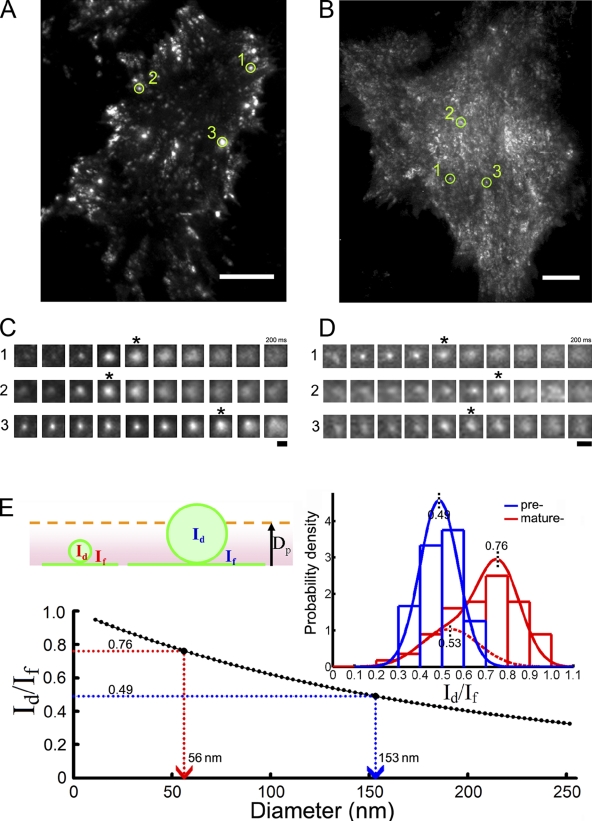
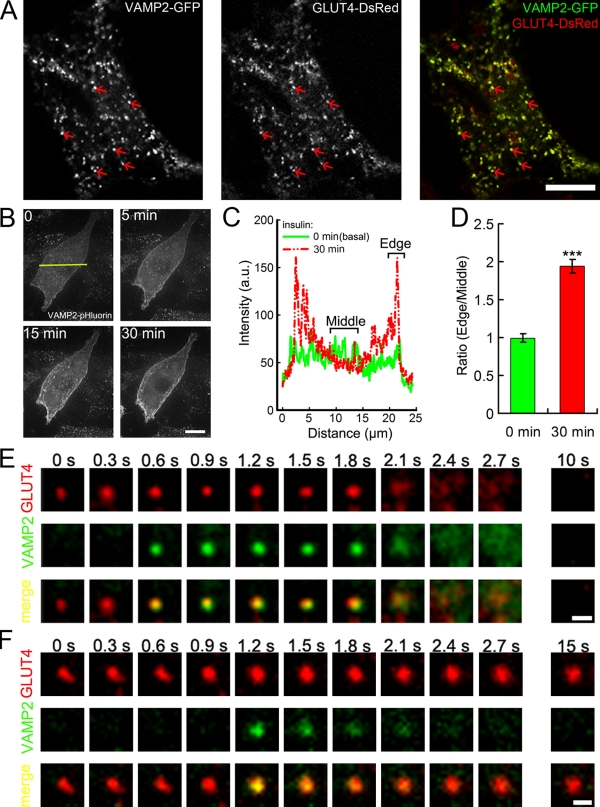
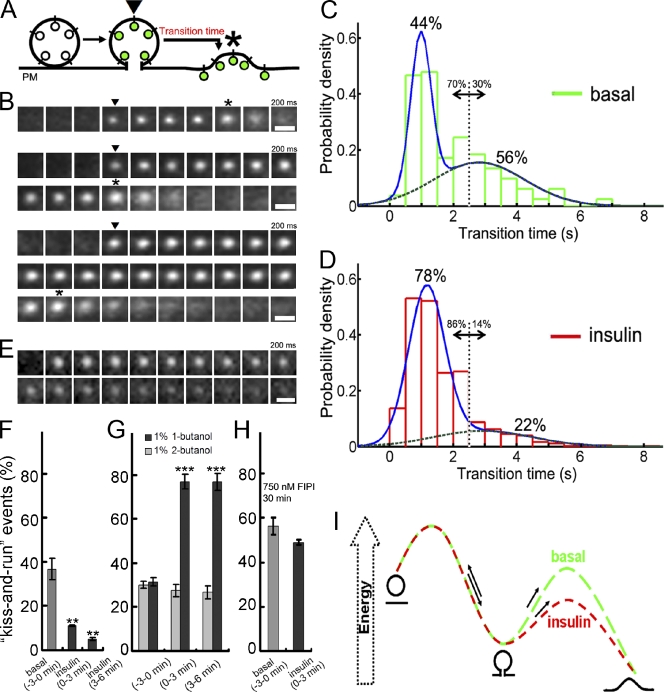
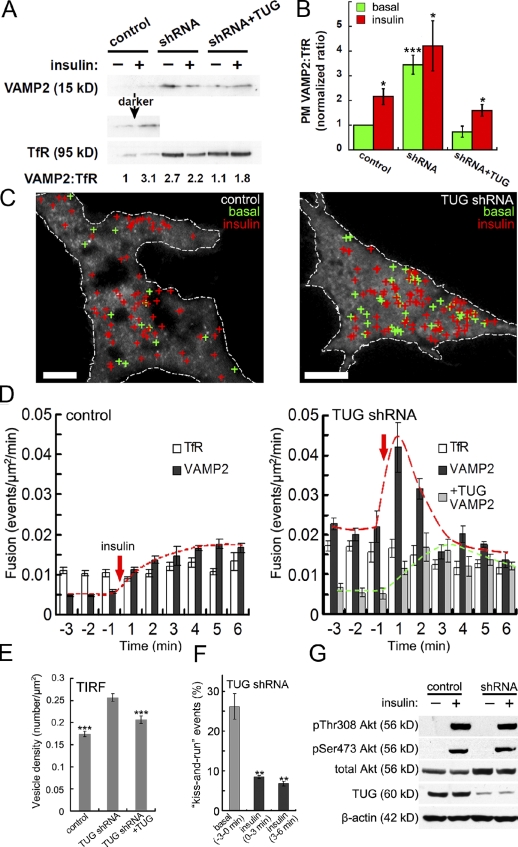
Insulin stimulates translocation of GLUT4 storage vesicles (GSVs) to the surface of adipocytes, but precisely where insulin acts is controversial. Here we quantify the size, dynamics, and frequency of single vesicle exocytosis in 3T3-L1 adipocytes. We use a new GSV reporter, VAMP2-pHluorin, and bypass insulin signaling by disrupting the GLUT4-retention protein TUG. Remarkably, in unstimulated TUG-depleted cells, the exocytic rate is similar to that in insulin-stimulated control cells. In TUG-depleted cells, insulin triggers a transient, twofold burst of exocytosis. Surprisingly, insulin promotes fusion pore expansion, blocked by acute perturbation of phospholipase D, which reflects both properties intrinsic to the mobilized vesicles and a novel regulatory site at the fusion pore itself. Prolonged stimulation causes cargo to switch from approximately 60 nm GSVs to larger exocytic vesicles characteristic of endosomes. Our results support a model whereby insulin promotes exocytic flux primarily by releasing an intracellular brake, but also by accelerating plasma membrane fusion and switching vesicle traffic between two distinct circuits.
Figures
Similar articles
-
GLUT4 Storage Vesicles: Specialized Organelles for Regulated Trafficking.Yale J Biol Med. 2019 Sep 20;92(3):453-470. eCollection 2019 Sep. Yale J Biol Med. 2019. PMID: 31543708 Free PMC article. Review.
-
Endoproteolytic cleavage of TUG protein regulates GLUT4 glucose transporter translocation.J Biol Chem. 2012 Jul 6;287(28):23932-47. doi: 10.1074/jbc.M112.339457. Epub 2012 May 18. J Biol Chem. 2012. PMID: 22610098 Free PMC article.
-
Acetylation of TUG protein promotes the accumulation of GLUT4 glucose transporters in an insulin-responsive intracellular compartment.J Biol Chem. 2015 Feb 13;290(7):4447-63. doi: 10.1074/jbc.M114.603977. Epub 2015 Jan 5. J Biol Chem. 2015. PMID: 25561724 Free PMC article.
-
Insulin-regulated Glut4 translocation: membrane protein trafficking with six distinctive steps.J Biol Chem. 2014 Jun 20;289(25):17280-98. doi: 10.1074/jbc.M114.555714. Epub 2014 Apr 28. J Biol Chem. 2014. PMID: 24778187 Free PMC article.
-
Spatiotemporal Regulators for Insulin-Stimulated GLUT4 Vesicle Exocytosis.J Diabetes Res. 2017;2017:1683678. doi: 10.1155/2017/1683678. Epub 2017 Apr 25. J Diabetes Res. 2017. PMID: 28529958 Free PMC article. Review.
Cited by
-
Sphingomyelin is sorted at the trans Golgi network into a distinct class of secretory vesicle.Proc Natl Acad Sci U S A. 2016 Jun 14;113(24):6677-82. doi: 10.1073/pnas.1602875113. Epub 2016 May 31. Proc Natl Acad Sci U S A. 2016. PMID: 27247384 Free PMC article.
-
Cholesterol accumulation increases insulin granule size and impairs membrane trafficking.Traffic. 2012 Nov;13(11):1466-80. doi: 10.1111/j.1600-0854.2012.01407.x. Epub 2012 Sep 13. Traffic. 2012. PMID: 22889194 Free PMC article.
-
Rab10 and myosin-Va mediate insulin-stimulated GLUT4 storage vesicle translocation in adipocytes.J Cell Biol. 2012 Aug 20;198(4):545-60. doi: 10.1083/jcb.201111091. J Cell Biol. 2012. PMID: 22908308 Free PMC article.
-
GLUT4 Storage Vesicles: Specialized Organelles for Regulated Trafficking.Yale J Biol Med. 2019 Sep 20;92(3):453-470. eCollection 2019 Sep. Yale J Biol Med. 2019. PMID: 31543708 Free PMC article. Review.
-
Novel ecto-tagged integrins reveal their trafficking in live cells.Nat Commun. 2017 Sep 18;8(1):570. doi: 10.1038/s41467-017-00646-w. Nat Commun. 2017. PMID: 28924207 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
