

Dual and opposing roles of the unfolded protein response regulated by IRE1alpha and XBP1 in proinsulin processing and insulin secretion
- PMID: 21555585
- PMCID: PMC3102350
- DOI: 10.1073/pnas.1105564108
Dual and opposing roles of the unfolded protein response regulated by IRE1alpha and XBP1 in proinsulin processing and insulin secretion
Abstract
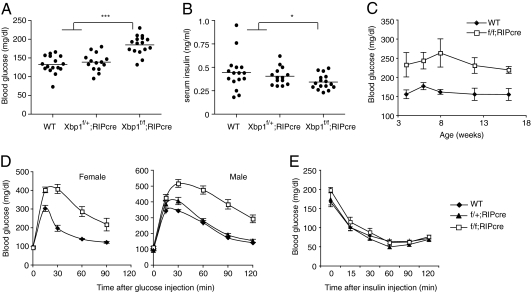
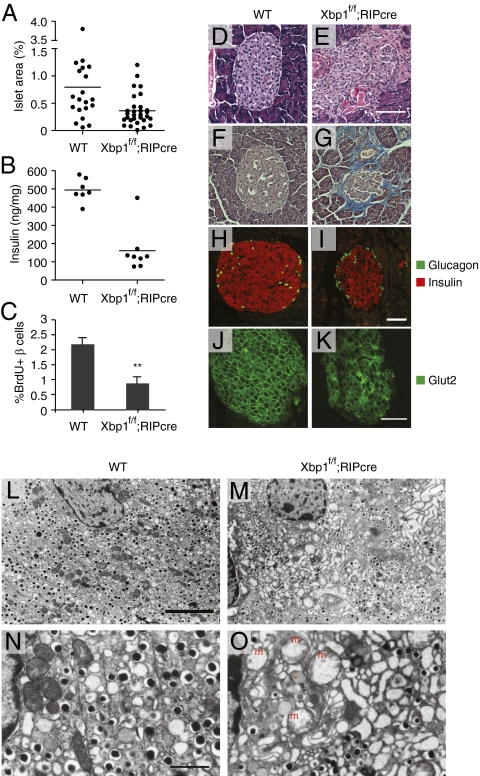
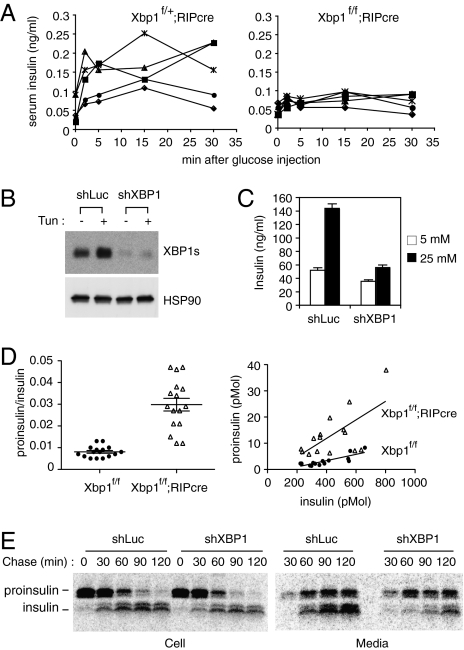
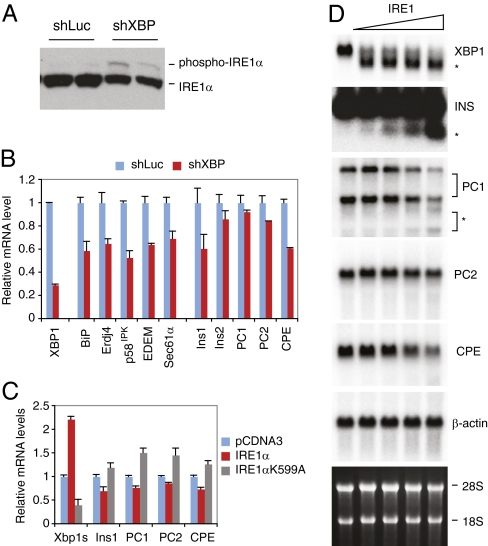
As a key regulator of the unfolded protein response, the transcription factor XBP1 activates genes in protein secretory pathways and is required for the development of certain secretory cells. To elucidate the function of XBP1 in pancreatic β-cells, we generated β-cell-specific XBP1 mutant mice. Xbp1(f/f);RIP-cre mice displayed modest hyperglycemia and glucose intolerance resulting from decreased insulin secretion from β-cells. Ablation of XBP1 markedly decreased the number of insulin granules in β-cells, impaired proinsulin processing, increased the serum proinsulin:insulin ratio, blunted glucose-stimulated insulin secretion, and inhibited cell proliferation. Notably, XBP1 deficiency not only compromised the endoplasmic reticulum stress response in β-cells but also caused constitutive hyperactivation of its upstream activator, IRE1α, which could degrade a subset of mRNAs encoding proinsulin-processing enzymes. Hence, the combined effects of XBP1 deficiency on the canonical unfolded protein response and its negative feedback activation of IRE1α caused β-cell dysfunction in XBP1 mutant mice. These results demonstrate that IRE1α has dual and opposing roles in β-cells, and that a precisely regulated feedback circuit involving IRE1α and its product XBP1s is required to achieve optimal insulin secretion and glucose control.
Conflict of interest statement
Conflict of interest statement: L.H.G. is a member of the board of directors of and holds equity in the Bristol Myers Squibb Corporation.
Figures
References
-
- Schröder M, Kaufman RJ. The mammalian unfolded protein response. Annu Rev Biochem. 2005;74:739–789. - PubMed
-
- Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol. 2007;8:519–529. - PubMed
-
- Kaufman RJ. Stress signaling from the lumen of the endoplasmic reticulum: Coordination of gene transcriptional and translational controls. Genes Dev. 1999;13:1211–1233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
