

miRNA-mediated silencing in hepatocytes can increase adaptive immune responses to adenovirus vector-delivered transgenic antigens
- PMID: 21556053
- PMCID: PMC3149177
- DOI: 10.1038/mt.2011.83
miRNA-mediated silencing in hepatocytes can increase adaptive immune responses to adenovirus vector-delivered transgenic antigens
Abstract
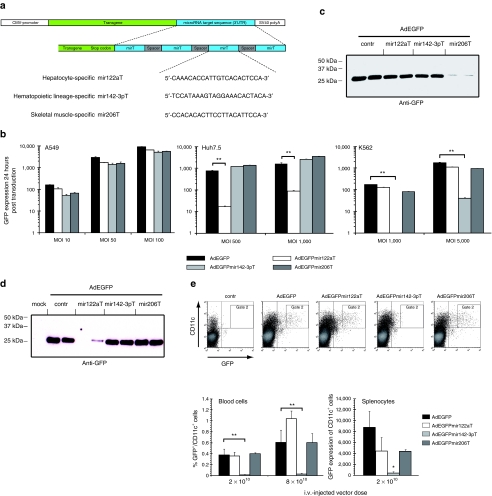
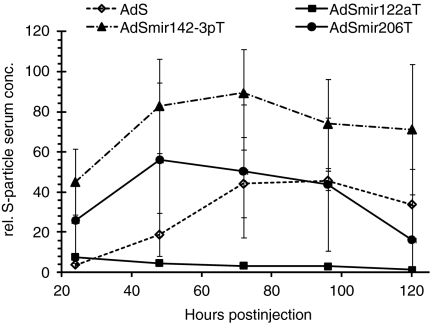
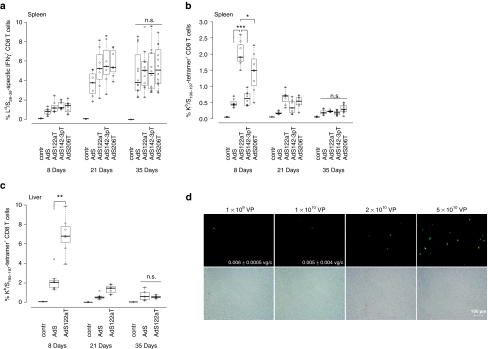
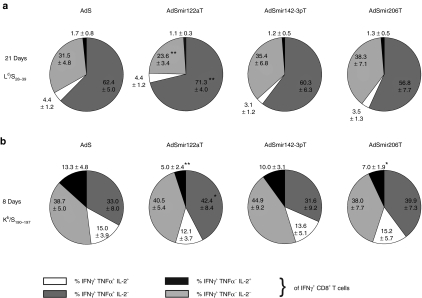
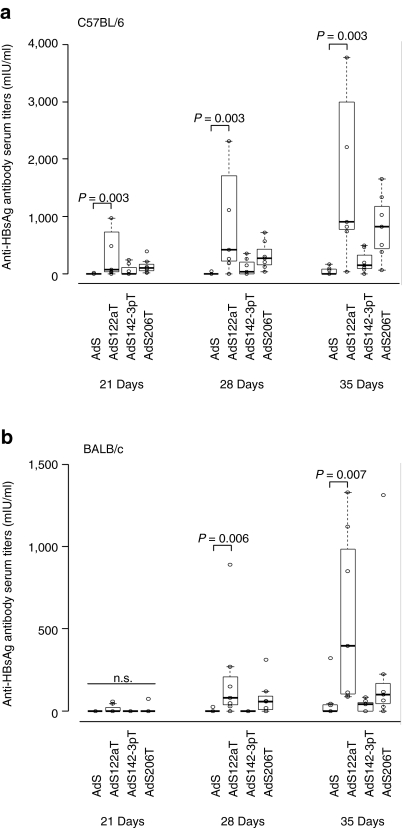
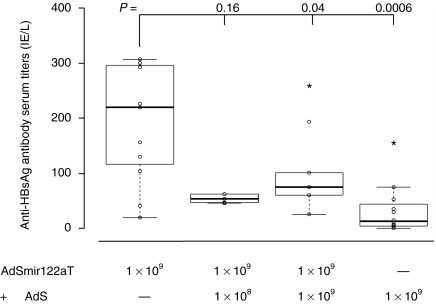
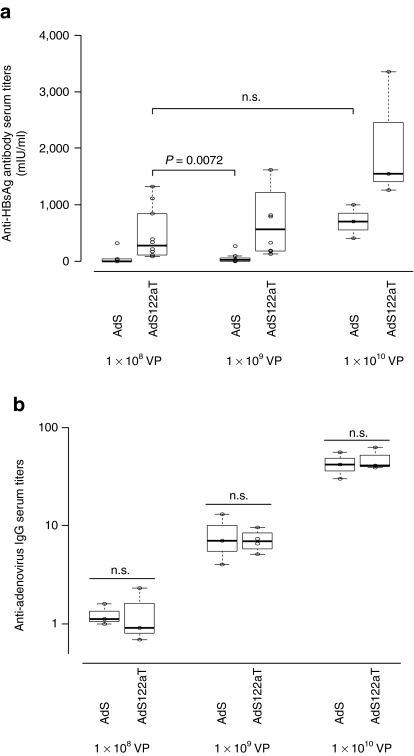
Adenovirus vectors based on human serotype 5 can induce potent CD8 T cell responses to vector-encoded transgenic antigens. However, the individual contribution of different cell types expressing antigen upon adenovirus vector injection to the generation of antigen-directed adaptive immune responses is poorly understood so far. We investigated the role of hepatocytes, skeletal muscle, and hematopoietic cells for the induction of cellular and humoral immune responses by miRNA-mediated tissue-specific silencing of antigen expression. Using hepatitis B small surface antigen (HBsAg) as the vector-encoded transgene we show that adenovirus vector dissemination from an intramuscular (i.m.) injection site into the liver followed by HBsAg expression in hepatocytes can limit early priming of CD8 T cells and the generation of anti-HBsAg antibody responses. However, hepatocyte-specific miRNA122a-mediated silencing of HBsAg expression overcame these limitations. Early clonal expansion of K(b)/S(190-197)-specific CD8 T cells was significantly enhanced and improved polyfunctionality of CD8 T cells was found. Furthermore, miRNA122a-mediated antigen silencing induced significantly higher anti-HBsAg antibody titers allowing an up to 100-fold vector dose reduction. These results indicate that miRNA-mediated regulation of antigen expression in the context of adenovirus vectors can significantly improve transgene product-directed immune responses. This finding could be of interest for future adenovirus vaccine vector development.
Figures
References
-
- Bergelson JM, Cunningham JA, Droguett G, Kurt-Jones EA, Krithivas A, Hong JS.et al. (1997Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5 Science 2751320–1323. - PubMed
-
- Wickham TJ, Mathias P, Cheresh DA., and, Nemerow GR. Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell. 1993;73:309–319. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
