

Genetic evidence points to an osteocalcin-independent influence of osteoblasts on energy metabolism
- PMID: 21557308
- PMCID: PMC3656486
- DOI: 10.1002/jbmr.417
Genetic evidence points to an osteocalcin-independent influence of osteoblasts on energy metabolism
Abstract
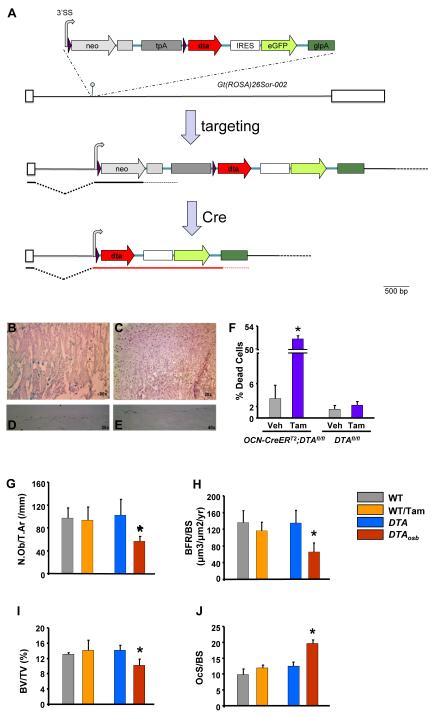
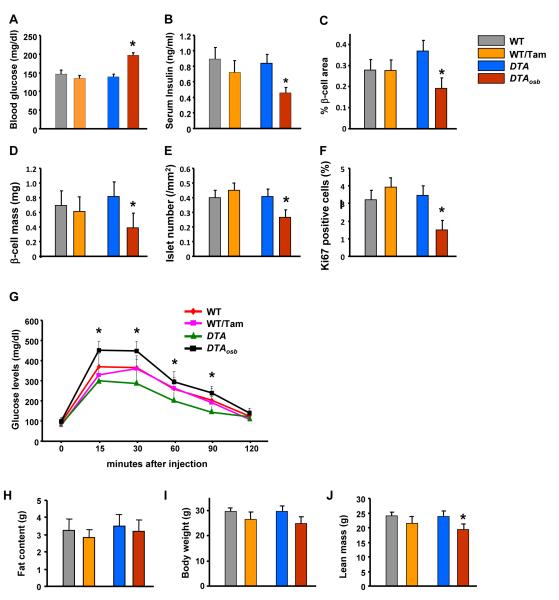
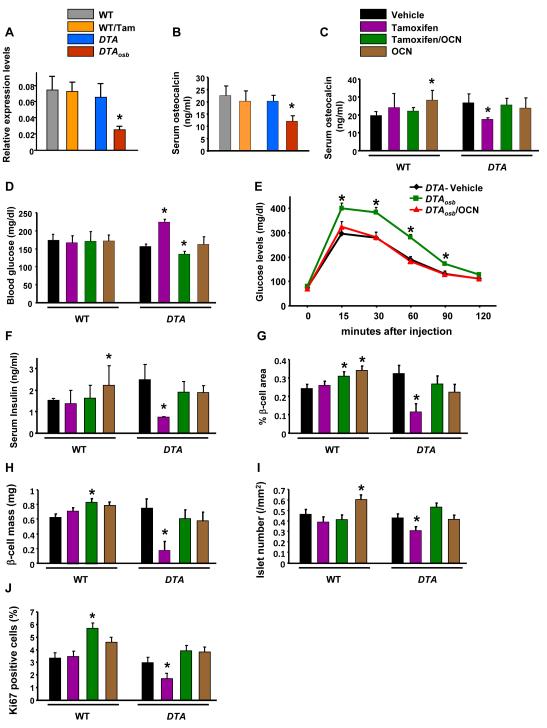
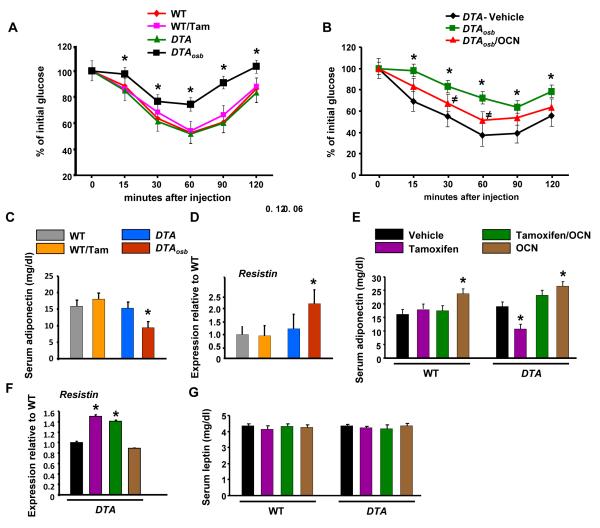
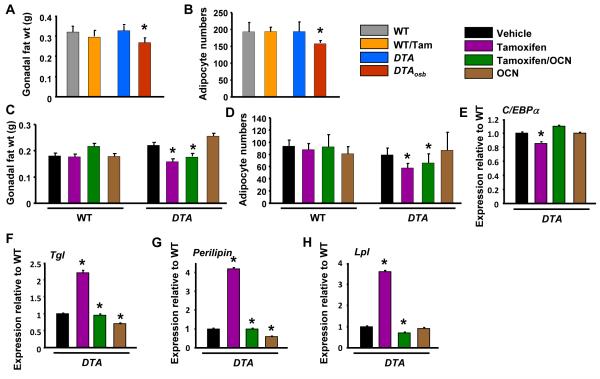
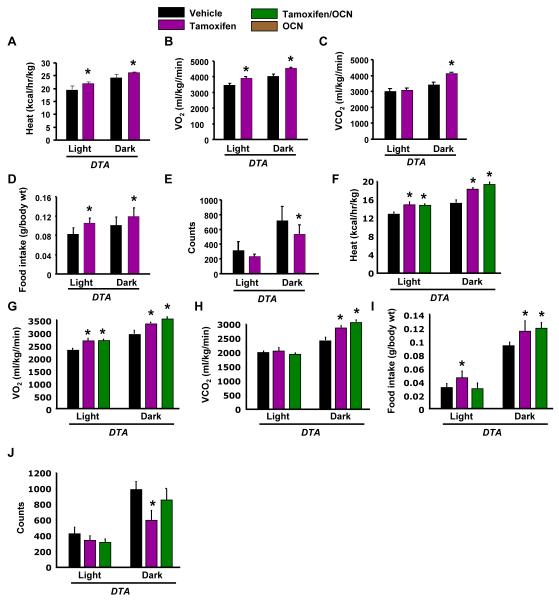
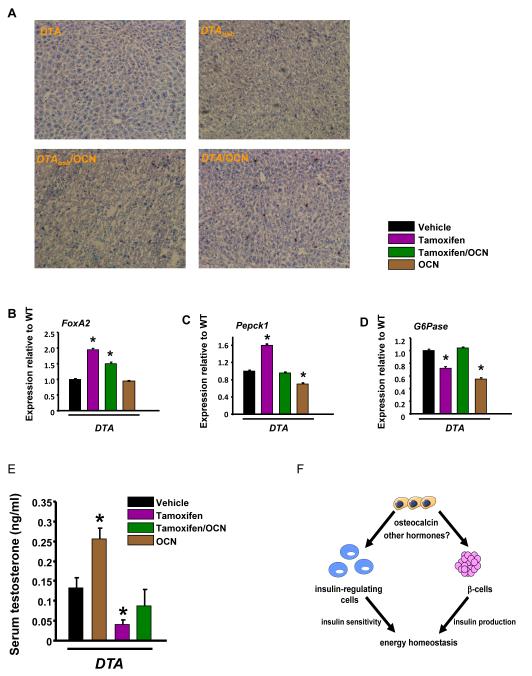
The skeleton has been shown recently to regulate glucose metabolism through an osteoblast-specific hormone, osteocalcin, which favors β-cell proliferation, insulin secretion, insulin sensitivity, and energy expenditure. An implication of this finding is that a decrease in osteoblast numbers would compromise glucose metabolism in an osteocalcin-dependent manner. To test this hypothesis, osteoblasts were inducibly ablated by cross-breeding transgenic mice expressing a tamoxifen-regulated Cre under the control of the osteocalcin promoter with mice in which an inactive form of the diphtheria toxin A chain was introduced into a ubiquitously expressed locus. Ablation of osteoblasts in adult mice profoundly affected glucose metabolism. In a manner similar to what is seen in the case of osteocalcin deficiency, a partial ablation of this cell population resulted in hypoinsulinemia, hyperglycemia, glucose intolerance, and decreased insulin sensitivity. However, and unlike what is seen in osteocalcin-deficient mice, osteoblast ablation also decreased gonadal fat and increased energy expenditure and the expression of resistin, an adipokine proposed to mediate insulin resistance. While administration of osteocalcin reversed (fully) the glucose intolerance and reinstated normal blood glucose and insulin levels, it only partially restored insulin sensitivity and did not affect the improved gonadal fat weight and energy expenditure in osteoblast-depleted mice. These observations not only strengthen the notion that osteoblasts are necessary for glucose homeostasis and energy expenditure but also suggest that in addition to osteocalcin, other osteoblast-derived hormones may contribute to the emerging function of the skeleton as a regulator of energy metabolism.
Copyright © 2011 American Society for Bone and Mineral Research.
Figures
References
-
- Kindblom JM, Ohlsson C, Ljunggren O, Karlsson MK, Tivesten A, Smith U, Mellstrom D. Plasma osteocalcin is inversely related to fat mass and plasma glucose in elderly Swedish men. J Bone Miner Res. 2009 May;24:785–791. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
