

Constitutive formation of an RXFP1-signalosome: a novel paradigm in GPCR function and regulation
- PMID: 21557732
- PMCID: PMC3372819
- DOI: 10.1111/j.1476-5381.2011.01470.x
Constitutive formation of an RXFP1-signalosome: a novel paradigm in GPCR function and regulation
Abstract
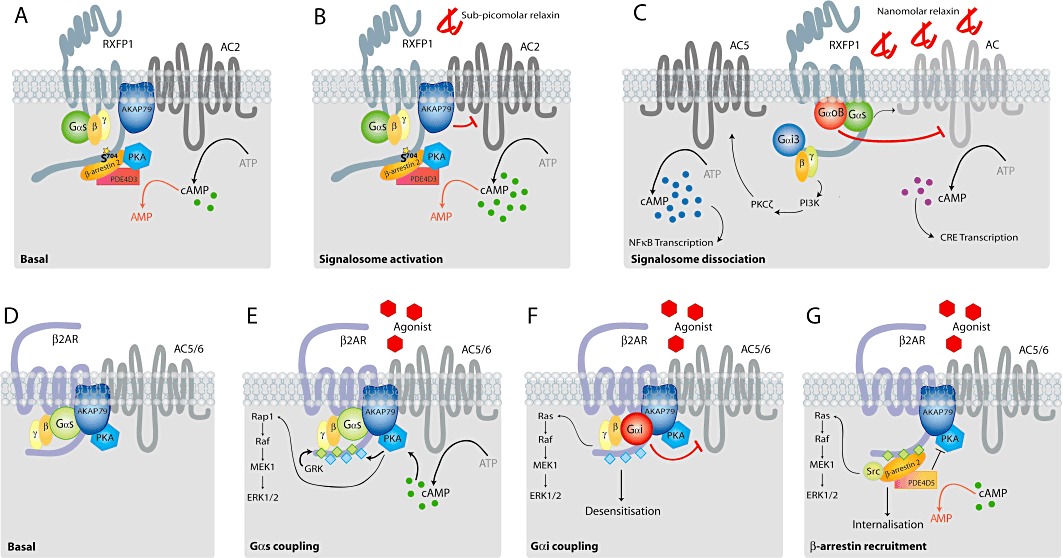
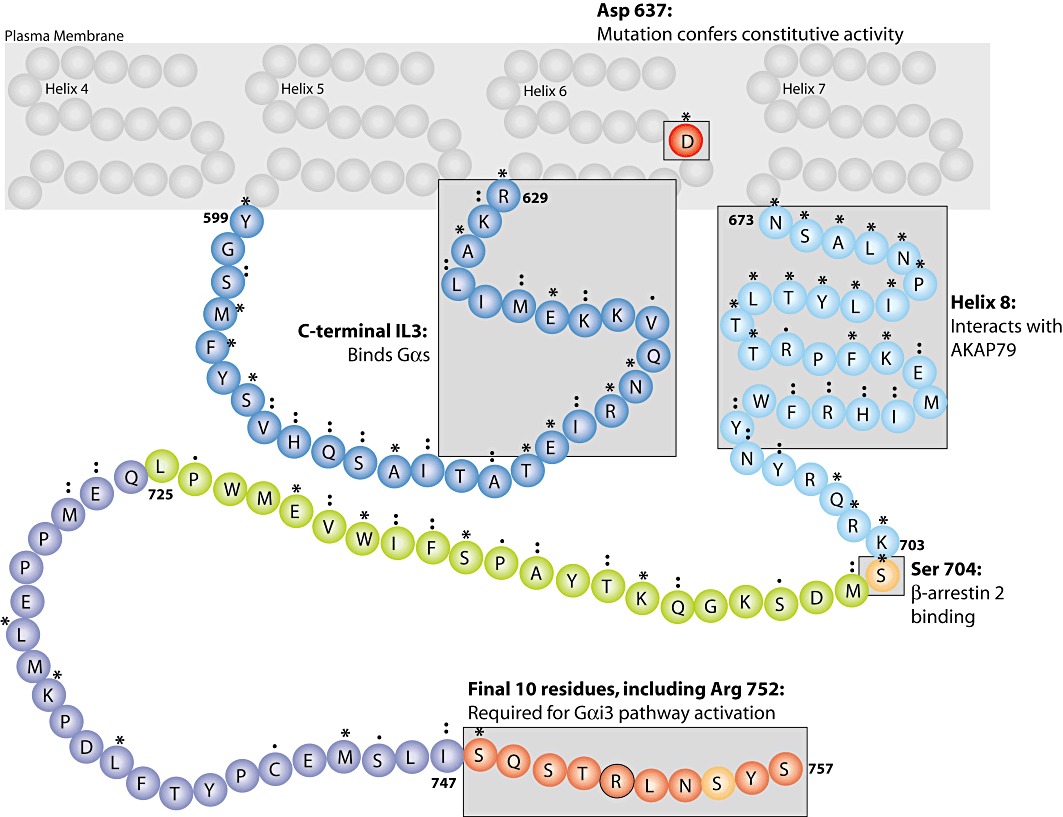
The classical second messenger cAMP is important in diverse physiological processes, where its spatial and temporal compartmentalization allows precise control over multiple cellular events. Within this context, G-protein-coupled receptors (GPCRs) govern specialized pools of cAMP, which are functionally specific for the unique cellular effects attributed to a particular system. The relaxin receptor, RXFP1, is a GPCR that exerts pleiotropic physiological effects including a potent anti-fibrotic response, increased cancer metastases, and has efficacy as a vasodilator in heart failure. On a cellular level, relaxin stimulation of RXFP1 results in the activation of multiple G-protein pathways affecting cAMP accumulation. Specificity and diversity in the cAMP signal generated by RXFP1 is controlled by differential G-protein coupling dependent upon the background of cellular expression, and cAMP compartmentalization. Further complexity in cAMP signalling results from the constitutive assembly of an RXFP1-signalosome, which specifically responds to low concentrations of relaxin, and activates a distinct cAMP pathway. The RXFP1-signalosome is a higher-order protein complex that facilitates receptor sensitivity to attomolar concentration of peptide, exhibits constitutive activity and dual coupling to G-proteins and β-arrestins and reveals a concentration-biased agonism mediated by relaxin. The specific and directed formation of GPCR-centered signalosomes allows an even greater spatial and temporal control of cAMP, thus rationalizing the considerable physiological scope of this ubiquitous second messenger.
© 2011 The Author. British Journal of Pharmacology © 2011 The British Pharmacological Society.
Figures
References
-
- Alleva DG, Kaser SB, Monroy MA, Fenton MJ, Beller DI. IL-15 functions as a potent autocrine regulator of macrophage proinflammatory cytokine production: evidence for differential receptor subunit utilization associated with stimulation or inhibition. J Immunol. 1997;159:2941–2951. - PubMed
-
- Anand-Ivell R, Heng K, Bartsch O, Ivell R. Relaxin signalling in THP-1 cells uses a novel phosphotyrosine-dependent pathway. Mol Cell Endocrinol. 2007;272:1–13. - PubMed
-
- Baillie GS, Scott JD, Houslay MD. Compartmentalisation of phosphodiesterases and protein kinase A: opposites attract. FEBS Lett. 2005;579:3264–3270. - PubMed
-
- Bartsch O, Bartlick B, Ivell R. Relaxin signalling links tyrosine phosphorylation to phosphodiesterase and adenylyl cyclase activity. Mol Hum Reprod. 2001;7:799–809. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
