

Structural requirements for the inhibition of calcium mobilization and mast cell activation by the pyrazole derivative BTP2
- PMID: 21558014
- PMCID: PMC3143500
- DOI: 10.1016/j.biocel.2011.04.016
Structural requirements for the inhibition of calcium mobilization and mast cell activation by the pyrazole derivative BTP2
Abstract
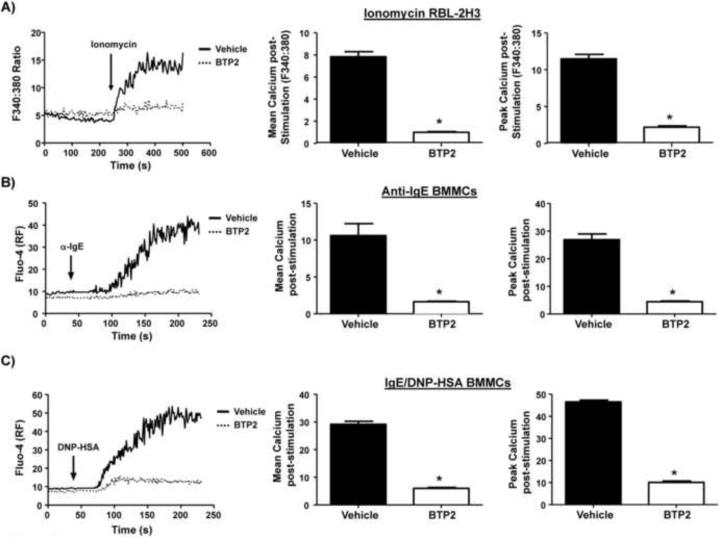
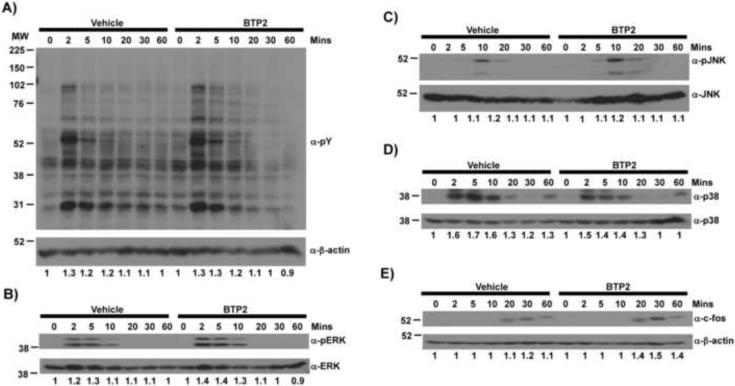
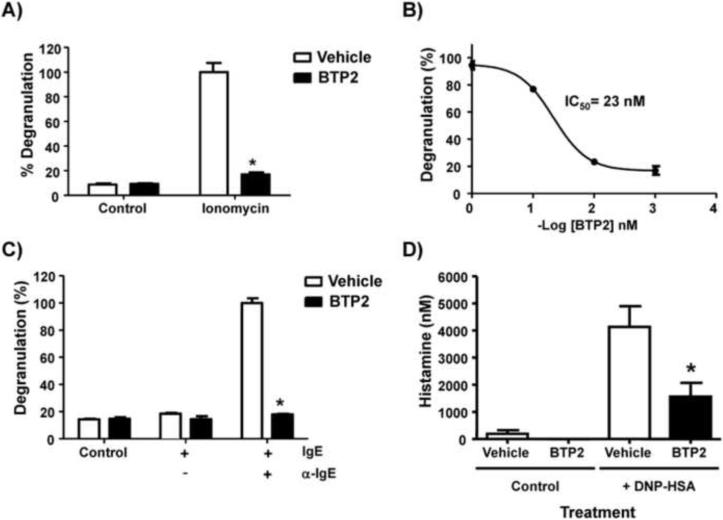
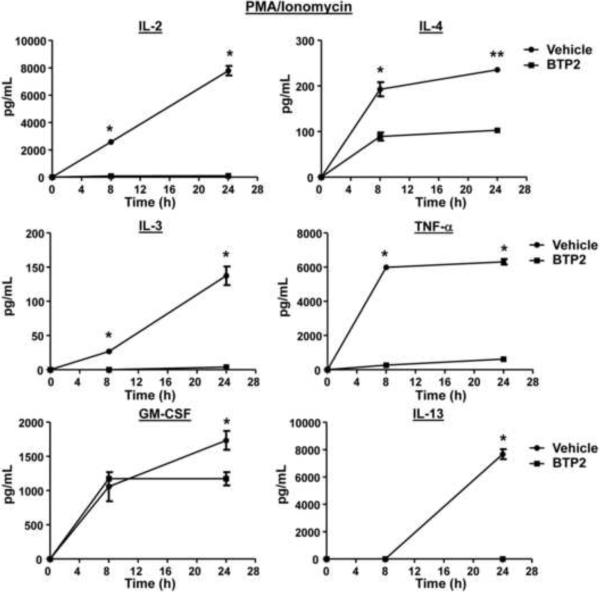
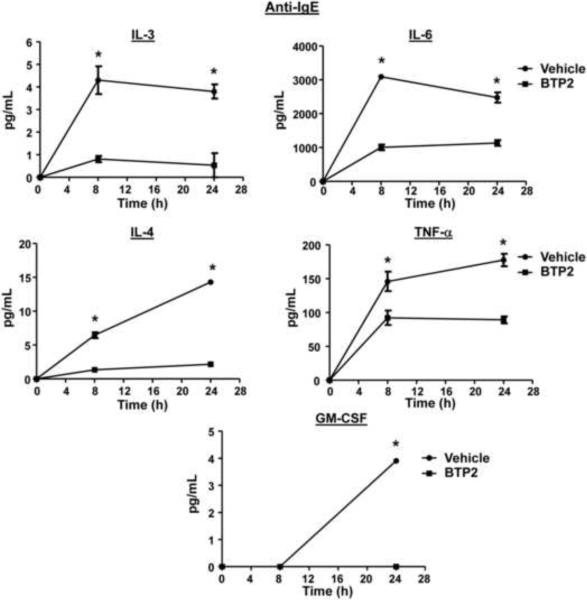
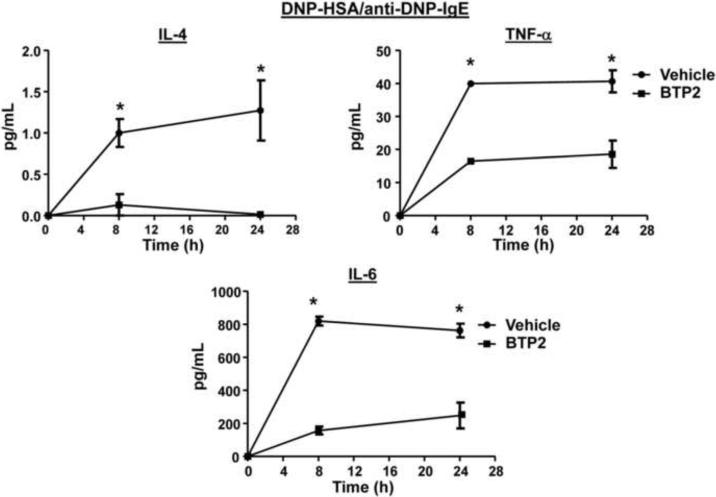
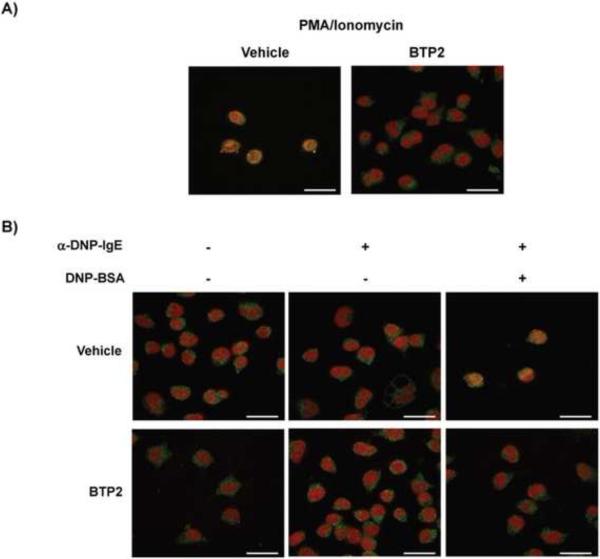
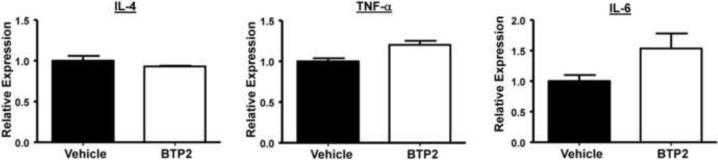
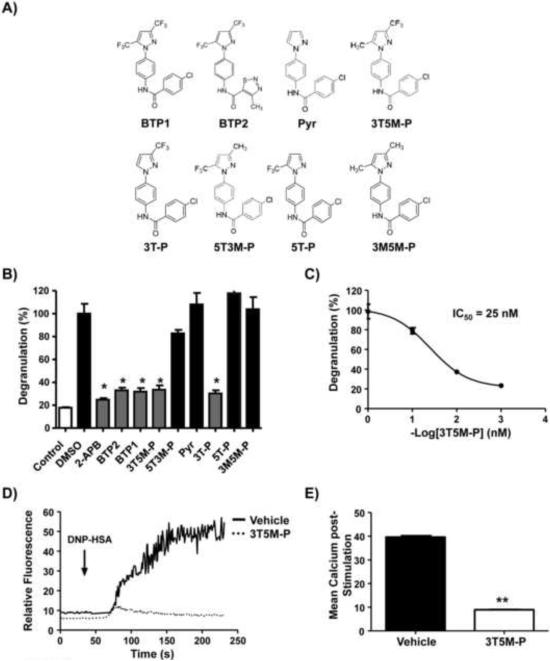
Mast cells play a critical role in the development of the allergic response. Upon activation by allergens and IgE via the high affinity receptor for IgE (FcɛRI), these cells release histamine and other functional mediators that initiate and propagate immediate hypersensitivity reactions. Mast cells also secrete cytokines that can regulate immune activity. These processes are controlled, in whole or part, by increases in intracellular Ca(2+) induced by the FcɛRI. We show here that N-(4-(3,5-bis(trifluoromethyl)-1H-pyrazol-1-yl)phenyl)-4-methyl-1,2,3-thiadiazole-5-carboxamide (BTP2), a pyrazole derivative, inhibits activation-induced Ca(2+) influx in the rat basophil cell line RBL-2H3 and in bone marrow-derived mast cells (BMMCs), without affecting global tyrosine phosphorylation of cellular proteins or phosphorylation of the mitogen-activated protein kinases Erk1/2, JNK and p38. BTP2 also inhibits activation-induced degranulation and secretion of interleukin (IL)-2, IL-3, IL-4, IL-6, IL-13, tumor necrosis factor (TNF)-α, and granulocyte macrophage-colony stimulating factor (GM-CSF) by BMMCs, which correlates with the inhibition of Nuclear Factor of Activated T cells (NFAT) translocation. In vivo, BTP2 inhibits antigen-induced histamine release. Structure-activity relationship analysis indicates that substitution at the C3 or C5 position of the pyrazole moiety on BTP2 (5-trifluoromethyl-3-methyl-pyrazole or 3-trifluoromethyl-5-methyl-pyrazole, respectively) affected its activity, with the trifluoromethyl group at the C3 position being critical to its activity. We conclude that BTP2 and related compounds may be potent modulators of mast cell responses and potentially useful for the treatment of symptoms of allergic inflammation.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Baba Y, Nishida K, Fujii Y, Hirano T, Hikida M, Kurosaki T. Essential function for the calcium sensor STIM1 in mast cell activation and anaphylactic responses. Nat Immunol. 2008;9:81–88. - PubMed
-
- Braun FJ, Aziz O, Putney JW., Jr. 2-aminoethoxydiphenyl borane activates a novel calcium-permeable cation channel. Mol Pharmacol. 2003;63:1304–1311. - PubMed
-
- De Boer M, Mordvinov V, Thomas M, Sanderson C. Role of nuclear factor of activated T cells (NFAT) in the expression of interleukin-5 and other cytokines involved in the regulation of hemopoetic cells. Int J Biochem Cell Biol. 1999;31:1221–1236. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
