

Lateral Connectivity in the Olfactory Bulb is Sparse and Segregated
- PMID: 21559072
- PMCID: PMC3084525
- DOI: 10.3389/fncir.2011.00005
Lateral Connectivity in the Olfactory Bulb is Sparse and Segregated
Abstract
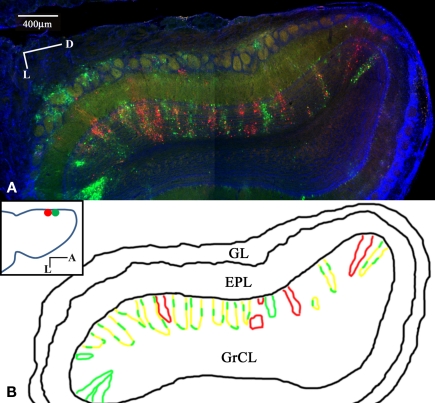
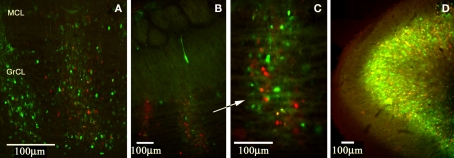
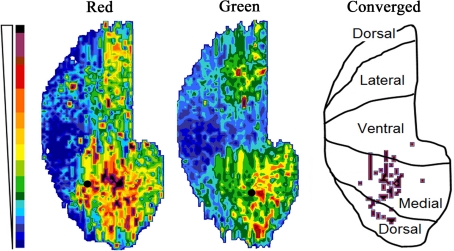
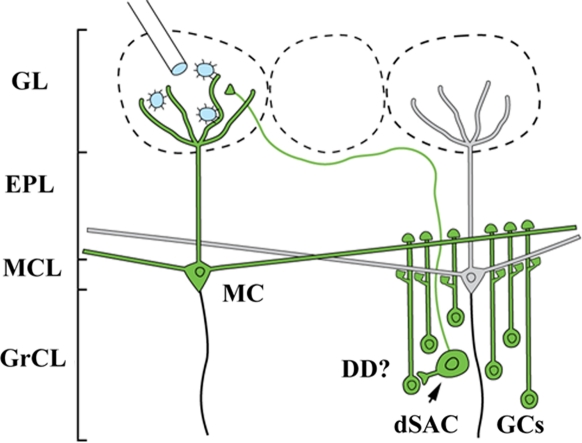
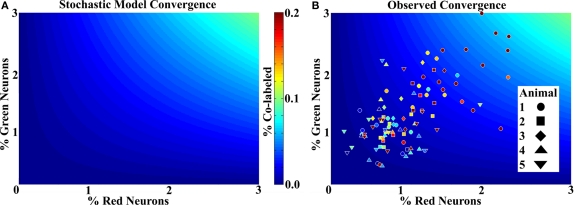
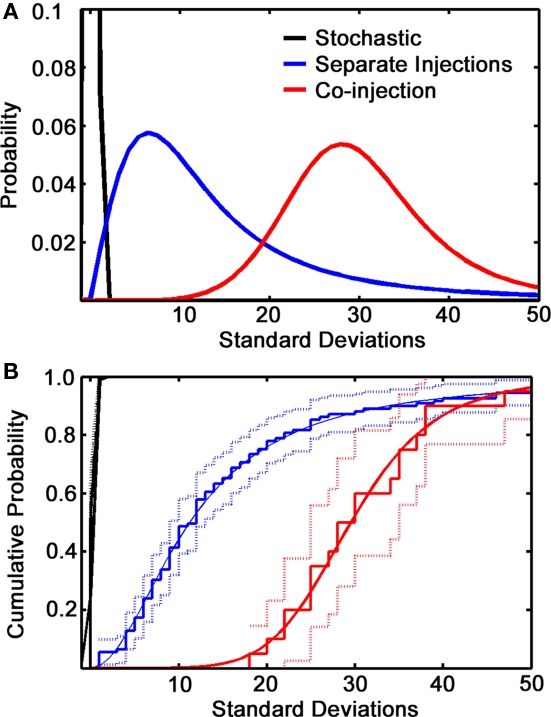
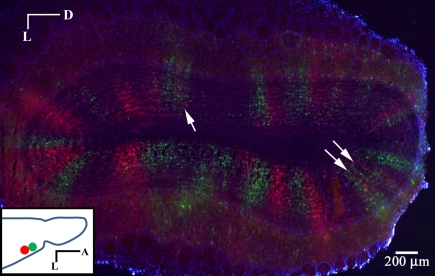
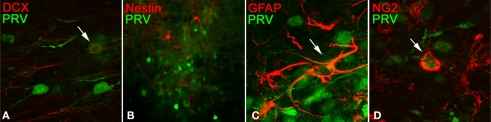
Lateral connections in the olfactory bulb were previously thought to be organized for center-surround inhibition. However, recent anatomical and physiological studies showed sparse and distributed interactions of inhibitory granule cells (GCs) which tended to be organized in columnar clusters. Little is known about how these distributed clusters are interconnected. In this study, we use transsynaptic tracing viruses bearing green or red fluorescent proteins to further elucidate mitral- and tufted-to-GC connectivity. Separate sites in the glomerular layer were injected with each virus. Columns with labeling from both viruses after transsynaptic spread show sparse red or green GCs which tended to be segregated. However, there was a higher incidence of co-labeled cells than chance would predict. Similar segregation of labeling is observed from dual injections into olfactory cortex. Collectively, these results suggest that neighboring mitral and tufted cells receive inhibitory inputs from segregated subsets of GCs, enabling inhibition of a center by specific and discontinuous lateral elements.
Keywords: circuits; connectivity; lateral inhibition; olfaction; olfactory bulb.
Figures
References
-
- Araneda R. C., Kini A. D., Firestein S. (2000). The molecular receptive range of an odorant receptor. Nat. Neurosci. 3, 1248–1255 - PubMed
-
- Card J. P., Enquist L. W. (1999). “Transneuronal circuit analysis with pseudorabies virus,” in Current Protocols in Neuroscience, ed. Crawley J. N. (New York, NY: J. Wiley; ), 1.5.1–1.5.28
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous