

Effects of connectivity and recurrent local disturbances on community structure and population density in experimental metacommunities
- PMID: 21559336
- PMCID: PMC3084878
- DOI: 10.1371/journal.pone.0019525
Effects of connectivity and recurrent local disturbances on community structure and population density in experimental metacommunities
Abstract
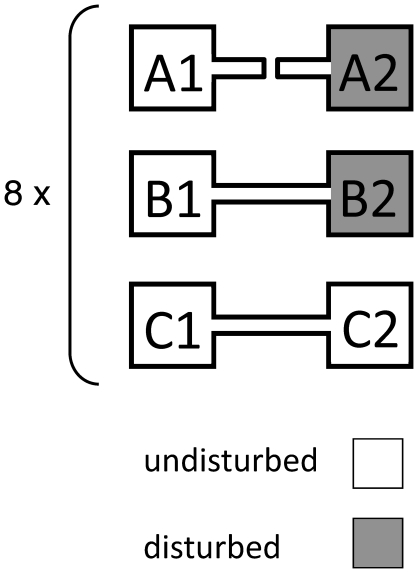
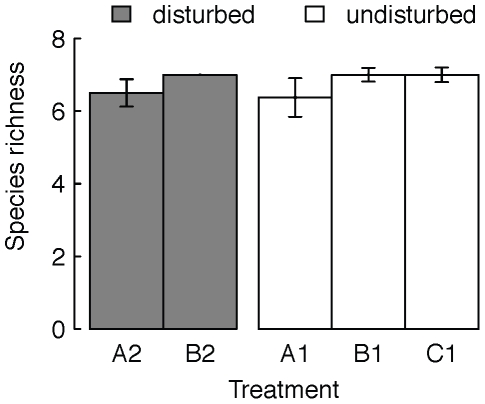
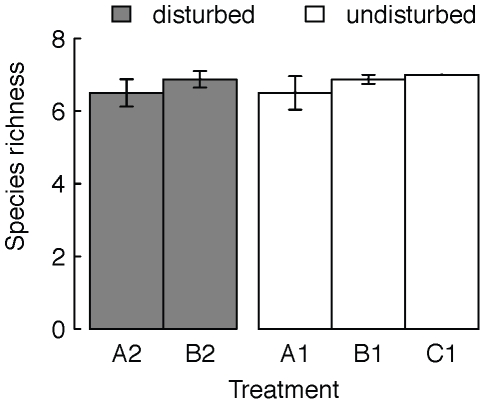
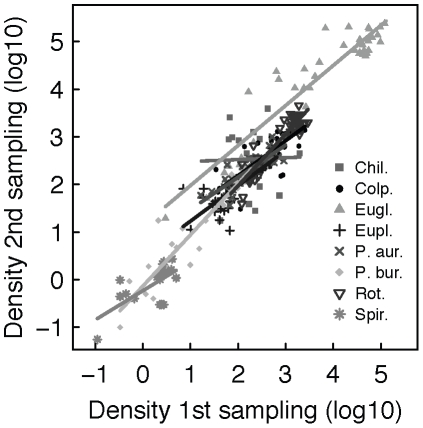
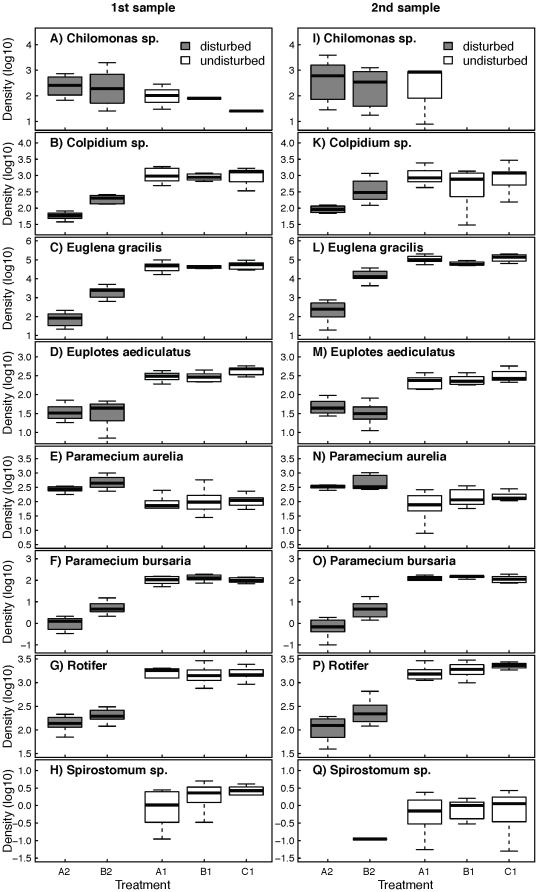
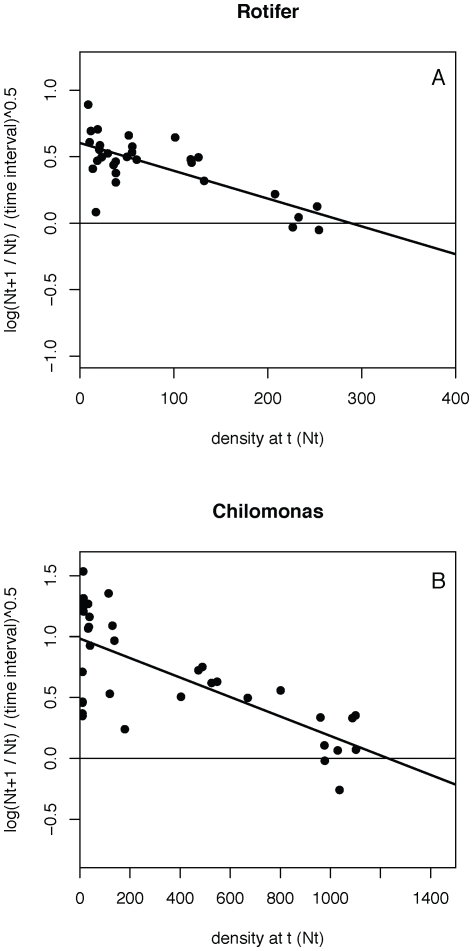
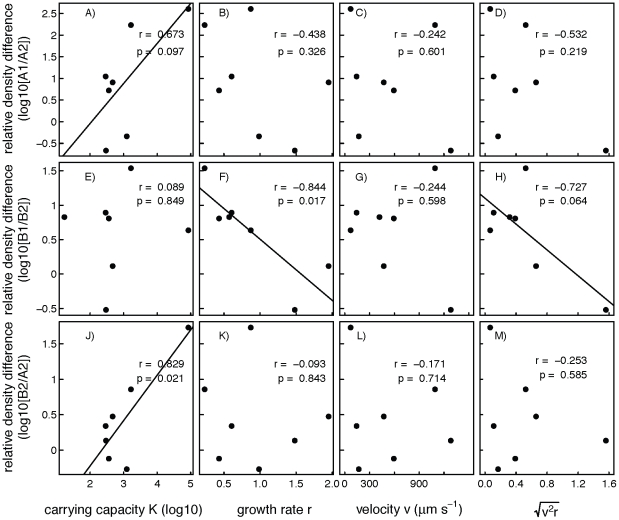
Metacommunity theory poses that the occurrence and abundance of species is a product of local factors, including disturbance, and regional factors, like dispersal among patches. While metacommunity ideas have been broadly tested there is relatively little work on metacommunities subject to disturbance. We focused on how localized disturbance and dispersal interact to determine species composition in metacommunities. Experiments conducted in simple two-patch habitats containing eight protozoa and rotifer species tested how dispersal altered community composition in both communities that were disturbed and communities that connected to refuge communities not subject to disturbance. While disturbance lowered population densities, in disturbed patches connected to undisturbed patches this was ameliorated by immigration. Furthermore, species with high dispersal abilities or growth rates showed the fastest post-disturbance recovery in presence of immigration. Connectivity helped to counteract the negative effect of disturbances on local populations, allowing mass-effect-driven dispersal of individuals from undisturbed to disturbed patches. In undisturbed patches, however, local population sizes were not significantly reduced by emigration. The absence of a cost of dispersal for undisturbed source populations is consistent with a lack of complex demography in our system, such as age- or sex-specific emigration. Our approach provides an improved way to separate components of population growth from organisms' movement in post-disturbance recovery of (meta)communities. Further studies are required in a variety of ecosystems to investigate the transient dynamics resulting from disturbance and dispersal.
Conflict of interest statement
Figures
References
-
- Holyoak M, Leibold MA, Holt RD, editors. Spatial dynamics and ecological communities. Chicago: The University of Chicago Press; 2005. Metacommunities.513
-
- Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, et al. The metacommunity concept: a framework for multi-scale community ecology. Ecology Letters. 2004;7:601–613.
-
- Gundersen G, Johannesen E, Andreassen HP, Ims RA. Source-sink dynamics: how sinks affect demography of sources. Ecology Letters. 2001;4:14–21.
-
- Holt RD. Ecology at the mesoscale: the influence of regional processes on local communities. In: Ricklefs RE, Schluter D, editors. Species Diversity in Ecological Communities: Historical and Geographical Perspectives. Chicago: University of Chicago Press; 1993. pp. 77–88.
-
- Amarasekare P. The role of density-dependent dispersal in source-sink dynamics. Journal of Theoretical Biology. 2004;226:159–168. - PubMed
