

How should prey animals respond to uncertain threats?
- PMID: 21559347
- PMCID: PMC3085230
- DOI: 10.3389/fncom.2011.00020
How should prey animals respond to uncertain threats?
Abstract
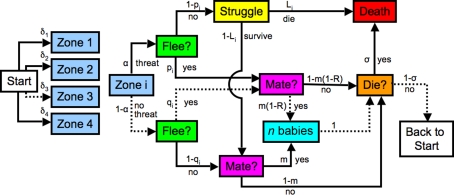
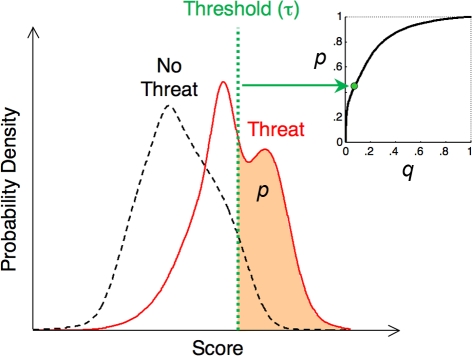
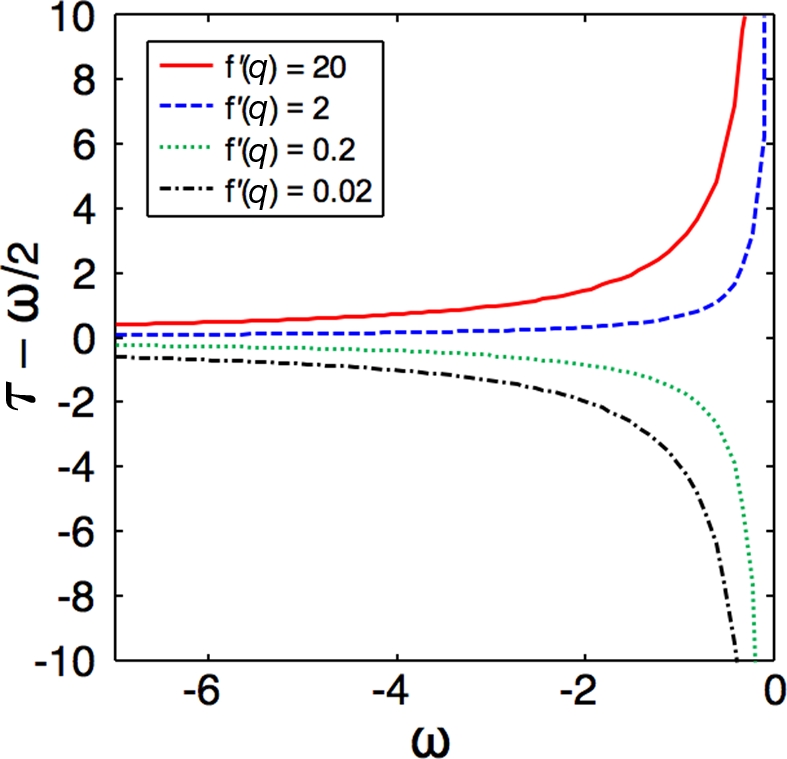
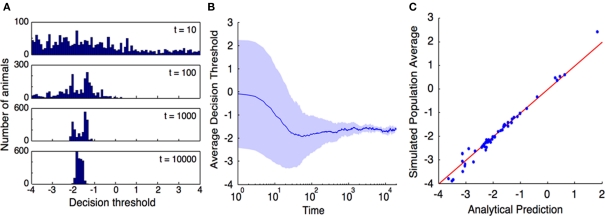
A prey animal surveying its environment must decide whether there is a dangerous predator present or not. If there is, it may flee. Flight has an associated cost, so the animal should not flee if there is no danger. However, the prey animal cannot know the state of its environment with certainty, and is thus bound to make some errors. We formulate a probabilistic automaton model of a prey animal's life and use it to compute the optimal escape decision strategy, subject to the animal's uncertainty. The uncertainty is a major factor in determining the decision strategy: only in the presence of uncertainty do economic factors (like mating opportunities lost due to flight) influence the decision. We performed computer simulations and found that in silico populations of animals subject to predation evolve to display the strategies predicted by our model, confirming our choice of objective function for our analytic calculations. To the best of our knowledge, this is the first theoretical study of escape decisions to incorporate the effects of uncertainty, and to demonstrate the correctness of the objective function used in the model.
Keywords: agent-based modeling; constrained optimization; decision making; escape decision; evolution; probabilistic automata; uncertainty.
Figures
References
-
- Benton T., Evans M. (1998). Measuring mate choice using correlation: the effect of sampling behaviour. Behav. Ecol. Sociobiol. 44, 91–9810.1007/s002650050520 - DOI
-
- Blumstein D. (2003). Flight-initiation distance in birds is dependent on intruder starting distance. J. Wildl. Manage. 64, 852–85710.2307/3802692 - DOI
LinkOut - more resources
Full Text Sources