

A novel tumour-suppressor function for the Notch pathway in myeloid leukaemia
- PMID: 21562564
- PMCID: PMC3093658
- DOI: 10.1038/nature09999
A novel tumour-suppressor function for the Notch pathway in myeloid leukaemia
Abstract
Notch signalling is a central regulator of differentiation in a variety of organisms and tissue types. Its activity is controlled by the multi-subunit γ-secretase (γSE) complex. Although Notch signalling can play both oncogenic and tumour-suppressor roles in solid tumours, in the haematopoietic system it is exclusively oncogenic, notably in T-cell acute lymphoblastic leukaemia, a disease characterized by Notch1-activating mutations. Here we identify novel somatic-inactivating Notch pathway mutations in a fraction of patients with chronic myelomonocytic leukaemia (CMML). Inactivation of Notch signalling in mouse haematopoietic stem cells (HSCs) results in an aberrant accumulation of granulocyte/monocyte progenitors (GMPs), extramedullary haematopoieisis and the induction of CMML-like disease. Transcriptome analysis revealed that Notch signalling regulates an extensive myelomonocytic-specific gene signature, through the direct suppression of gene transcription by the Notch target Hes1. Our studies identify a novel role for Notch signalling during early haematopoietic stem cell differentiation and suggest that the Notch pathway can play both tumour-promoting and -suppressive roles within the same tissue.
Figures
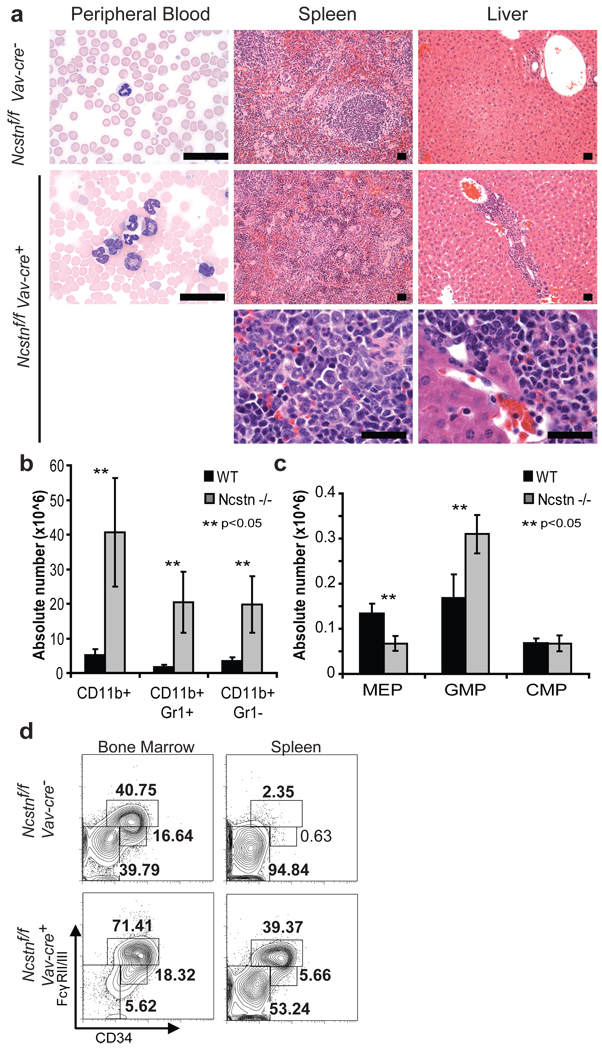
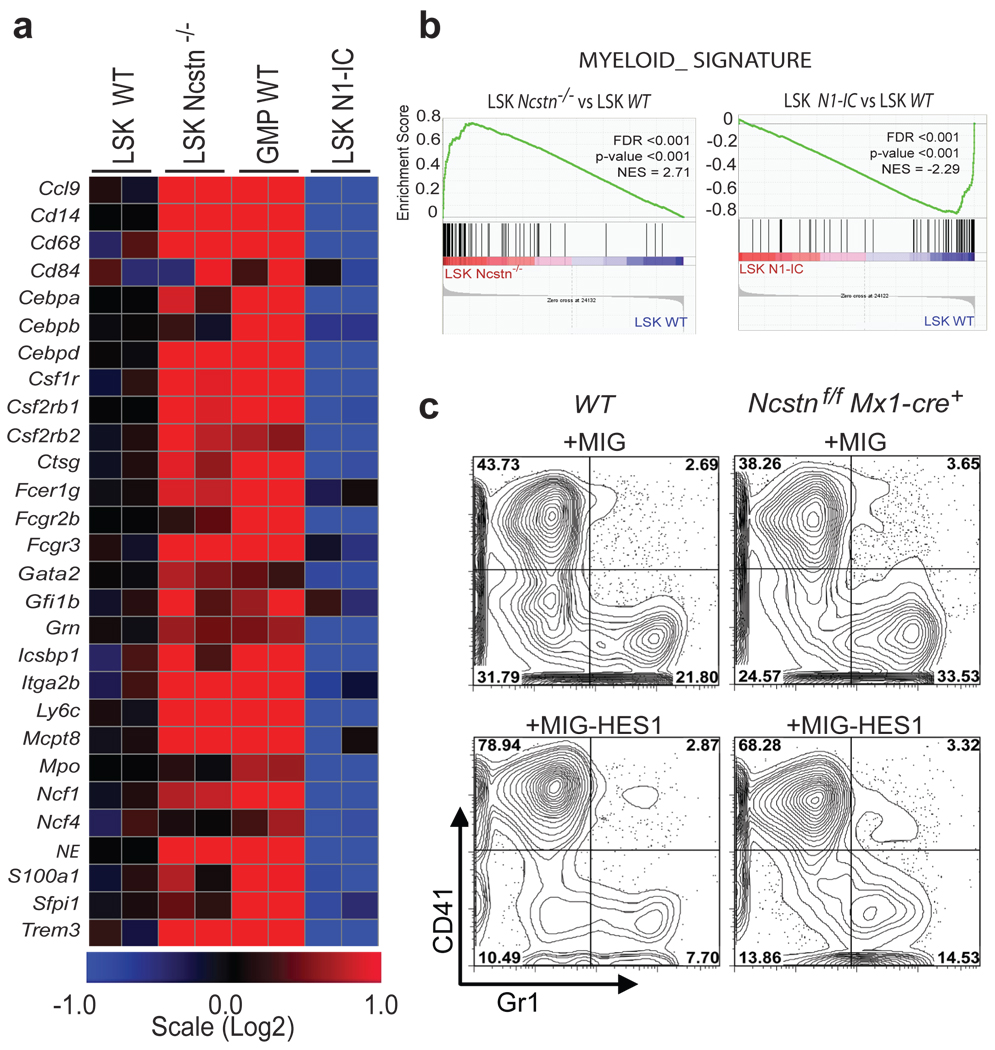
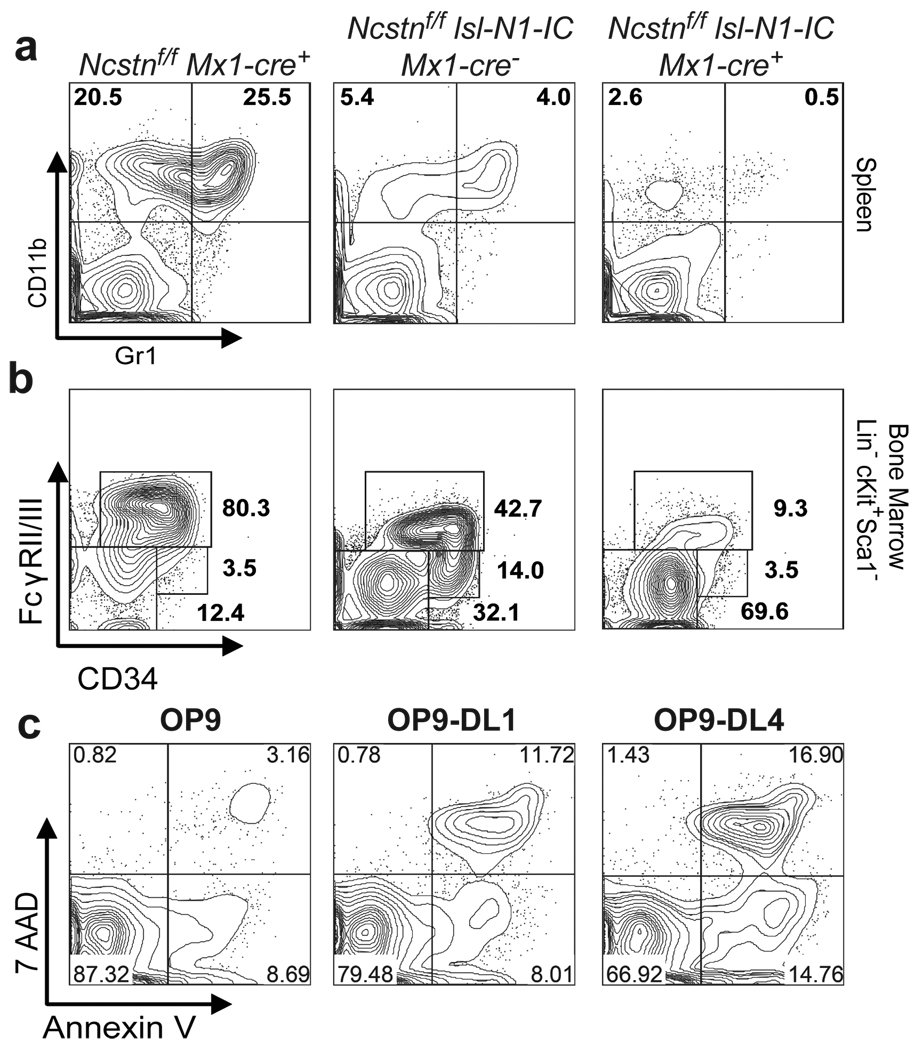
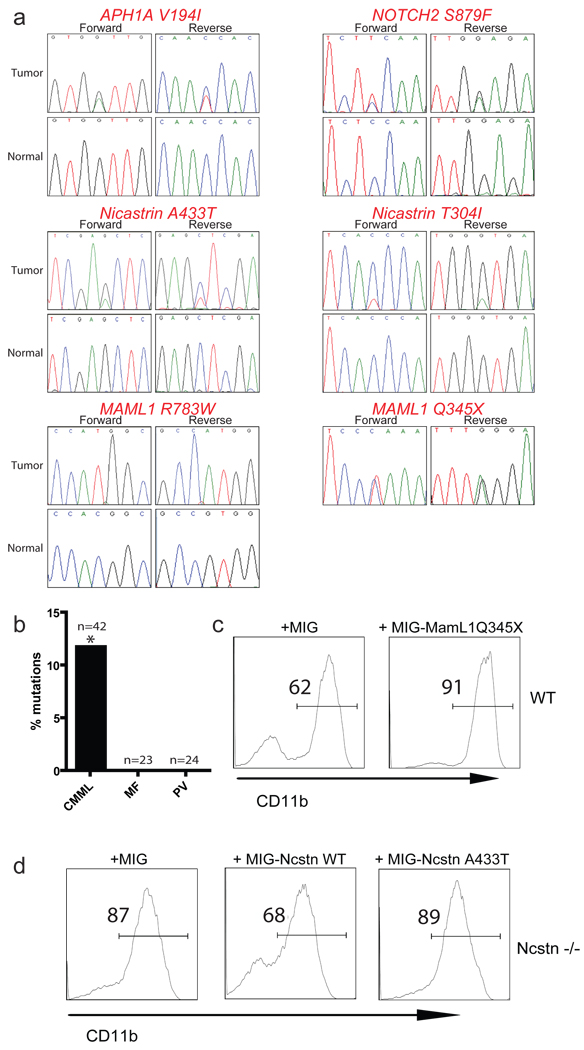
Comment in
-
Cancer: The flipside of Notch.Nature. 2011 May 12;473(7346):159-60. doi: 10.1038/473159a. Nature. 2011. PMID: 21562551 No abstract available.
-
Leukaemia: Notch has commitment issues.Nat Rev Cancer. 2011 Jun;11(6):385. doi: 10.1038/nrc3071. Nat Rev Cancer. 2011. PMID: 21606933 No abstract available.
-
Taking HSCs down a Notch in leukemia.Cell Stem Cell. 2011 Jun 3;8(6):602-3. doi: 10.1016/j.stem.2011.04.010. Cell Stem Cell. 2011. PMID: 21624800
References
-
- Hurlbut GD, Kankel MW, Lake RJ, Artavanis-Tsakonas S. Crossing paths with Notch in the hyper-network. Curr Opin Cell Biol. 2007;19:166–175. doi:S0955-0674(07)00026-9 [pii] 10.1016/j.ceb.2007.02.012. - PubMed
-
- De Strooper B. Nicastrin: gatekeeper of the gamma-secretase complex. Cell. 2005;122:318–320. doi:S0092-8674(05)00751-8 [pii] 10.1016/j.cell.2005.07.021. - PubMed
-
- Aifantis I, Raetz E, Buonamici S. Molecular pathogenesis of T-cell leukaemia and lymphoma. Nature reviews. 2008;8:380–390. - PubMed
-
- Zuniga-Pflucker JC. T-cell development made simple. Nature reviews. 2004;4:67–72. - PubMed
Methods References
-
- Yang X, et al. Notch activation induces apoptosis in neural progenitor cells through a p53-dependent pathway. Dev Biol. 2004;269:81–94. doi:10.1016/j.ydbio.2004.01.014 S0012160604000636 [pii] - PubMed
-
- Saito T, et al. Notch2 is preferentially expressed in mature B cells and indispensable for marginal zone B lineage development. Immunity. 2003;18:675–685. - PubMed
-
- Mitchell KJ, et al. Functional analysis of secreted and transmembrane proteins critical to mouse development. Nat Genet. 2001;28:241–249. - PubMed
-
- Leighton PA, et al. Defining brain wiring patterns and mechanisms through gene trapping in mice. Nature. 2001;410:174–179. - PubMed
-
- Rodriguez CI, et al. High-efficiency deleter mice show that FLPe is an alternative to Cre-loxP. Nat Genet. 2000;25:139–140. doi:10.1038/75973. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
- 1P01CA97403/CA/NCI NIH HHS/United States
- R01 CA133379/CA/NCI NIH HHS/United States
- R01CA133379/CA/NCI NIH HHS/United States
- R21CA141399/CA/NCI NIH HHS/United States
- R01 CA149655/CA/NCI NIH HHS/United States
- R01 CA105129/CA/NCI NIH HHS/United States
- R01CA1328234/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R21 CA141399/CA/NCI NIH HHS/United States
- R01CA149655/CA/NCI NIH HHS/United States
- U54 CA143798/CA/NCI NIH HHS/United States
- P01 CA097403/CA/NCI NIH HHS/United States
- U54CA143798/CA/NCI NIH HHS/United States
- R01CA105129/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
