

Site-directed spin labeling electron paramagnetic resonance study of the ORF1 protein from a mouse L1 retrotransposon
- PMID: 21563223
- PMCID: PMC3149196
- DOI: 10.1002/pro.651
Site-directed spin labeling electron paramagnetic resonance study of the ORF1 protein from a mouse L1 retrotransposon
Abstract
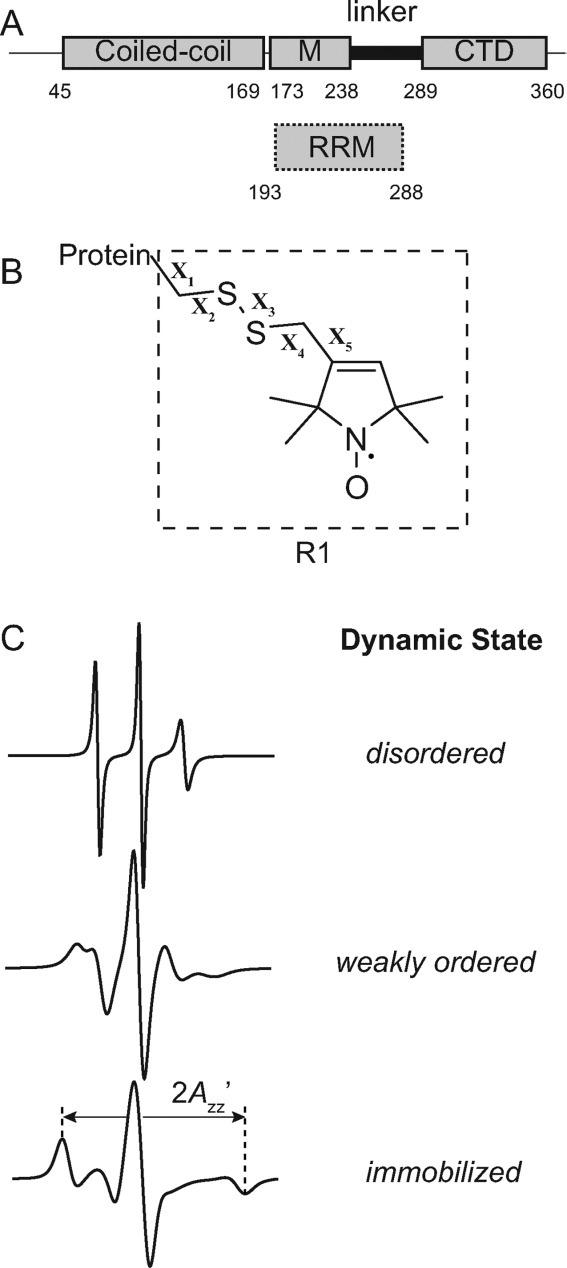
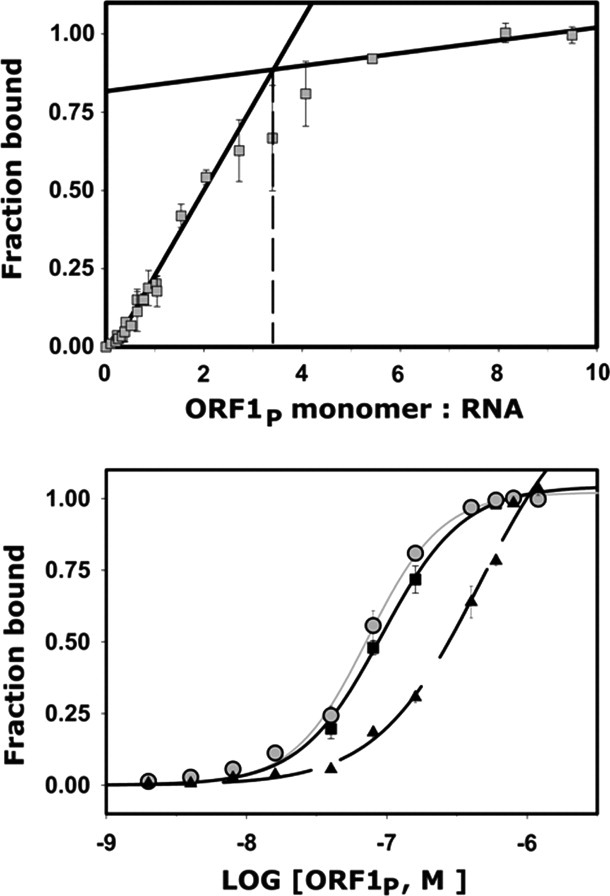
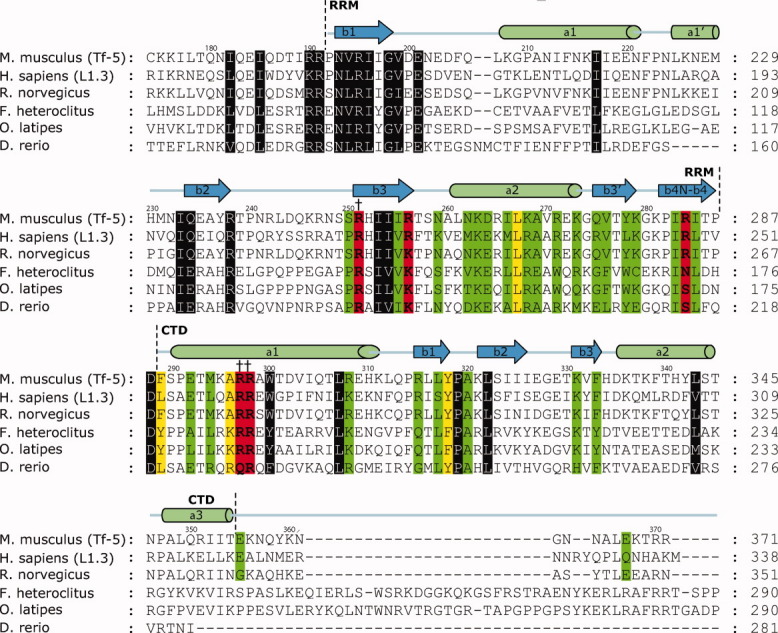
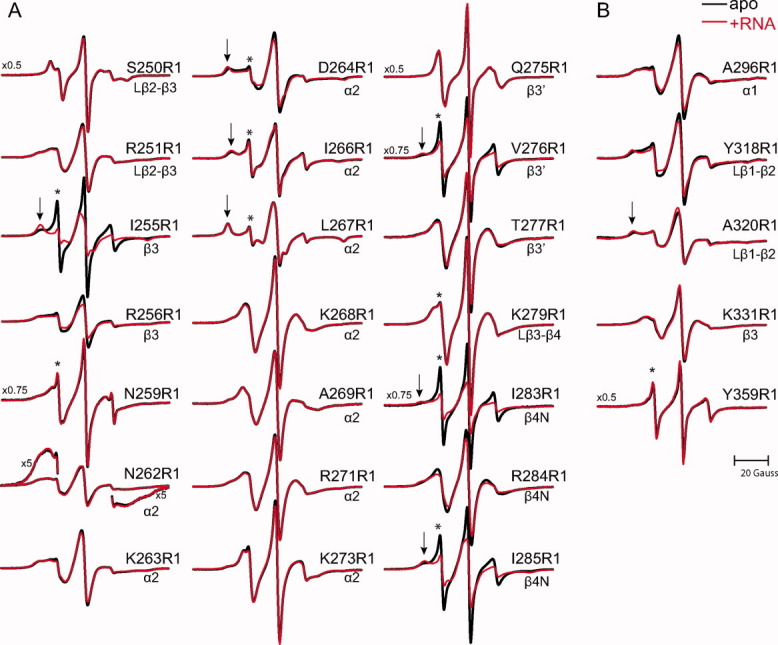
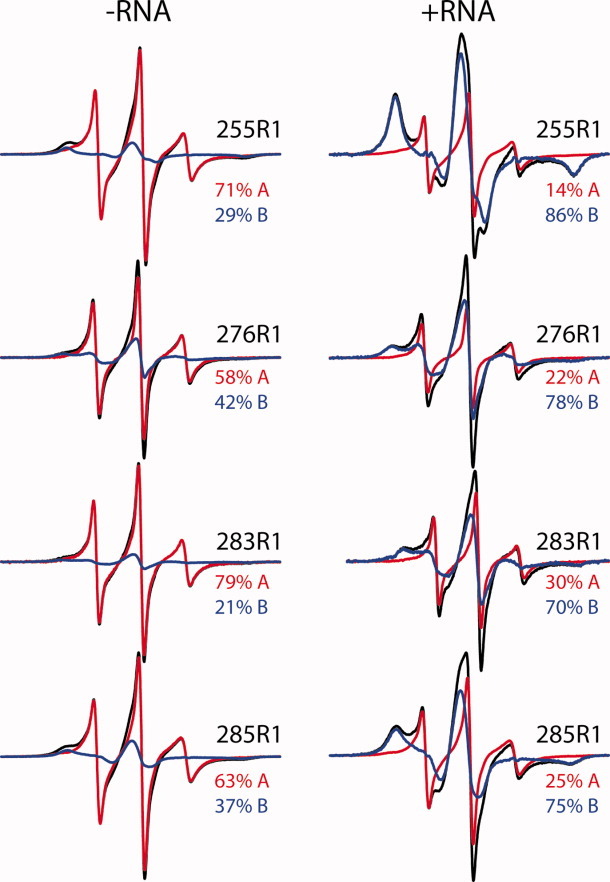
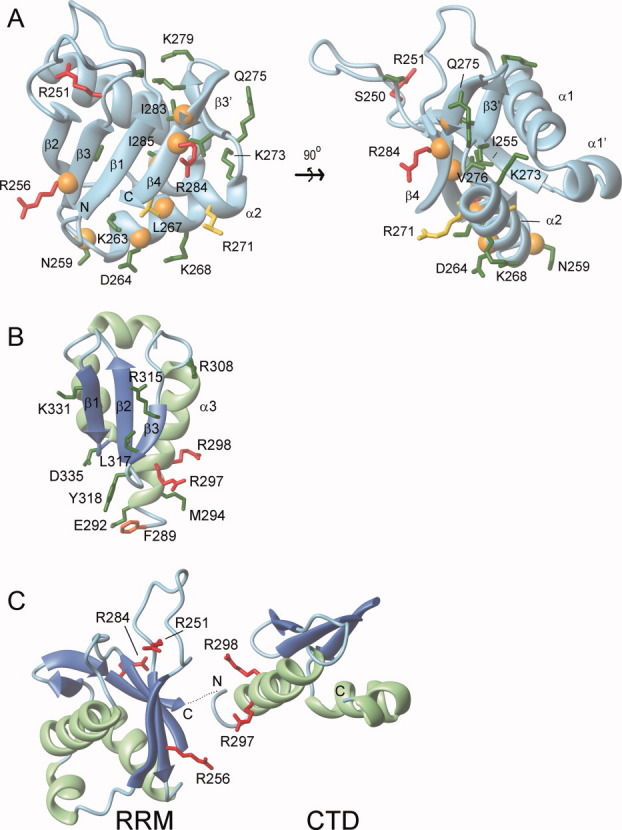
Long interspersed nuclear element-1 is a highly abundant mammalian retrotransposon that comprises 17% of the human genome. L1 retrotransposition requires the protein encoded by open reading frame-1 (ORF1p), which binds single-stranded RNA with high affinity and functions as a nucleic acid chaperone. ORF1p has been shown to adopt a homo-trimeric, asymmetric dumbbell-shaped structure. However, its atomic-level structure and mechanism of RNA binding remains poorly understood. Here, we report the results of a site-directed spin labeling electron paramagnetic resonance (SDSL-EPR) study of 27 residues within the RNA binding region of the full-length protein. The EPR data are compatible with the large RNA binding lobe of ORF1p containing a RNA recognition motif (RRM) domain and a carboxyl-terminal domain (CTD) that are predicted from crystallographic and NMR studies of smaller fragments of the protein. Interestingly, the EPR data indicate that residues in strands β3 and β4 of the RRM are structurally unstable, compatible with the previously observed sensitivity of this region to proteolysis. Affinity measurements and RNA-dependent EPR spectral changes map the RNA binding site on ORF1p to residues located in strands β3 and β4 of the RRM domain and to helix α1 of the CTD. Complementary in vivo studies also identify residues within the RRM domain that are required for retrotransposition. We propose that in the context of the full-length trimeric protein these distinct surfaces are positioned adjacent to one another providing a continuous surface that may interact with nucleic acids.
Copyright © 2011 The Protein Society.
Figures
References
-
- Goodier JL, Kazazian HH., Jr Retrotransposons revisited: the restraint and rehabilitation of parasites. Cell. 2008;135:23–35. - PubMed
-
- Babushok DV, Kazazian HH., Jr. Progress in understanding the biology of the human mutagen LINE-1. Hum Mutat. 2007;28:527–539. - PubMed
-
- Han JS, Boeke JD. LINE-1 retrotransposons: modulators of quantity and quality of mammalian gene expression? Bioessays. 2005;27:775–784. - PubMed
-
- Kazazian HH, Jr, Moran JV. The impact of L1 retrotransposons on the human genome. Nat Genet. 1998;19:19–24. - PubMed
-
- Ostertag EM, Kazazian HH. Biology of mammalian L1 retrotransposons. Annu Rev Genet. 2001;35:501–538. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
