

From an executive network to executive control: a computational model of the n-back task
- PMID: 21563882
- PMCID: PMC3269304
- DOI: 10.1162/jocn_a_00047
From an executive network to executive control: a computational model of the n-back task
Abstract
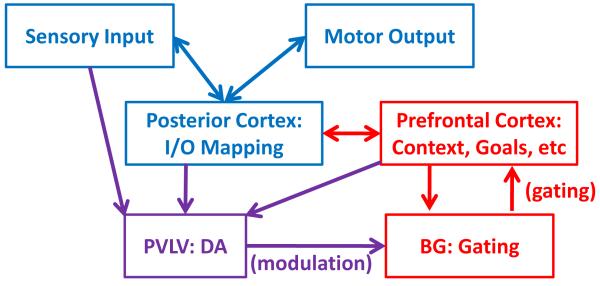
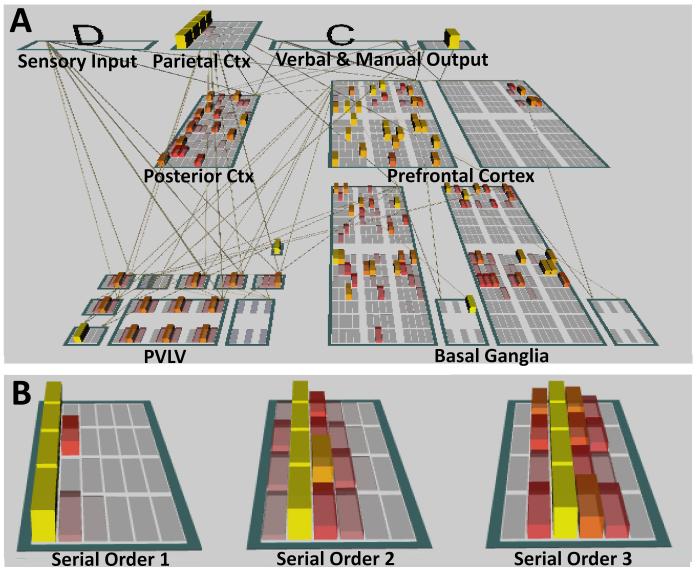
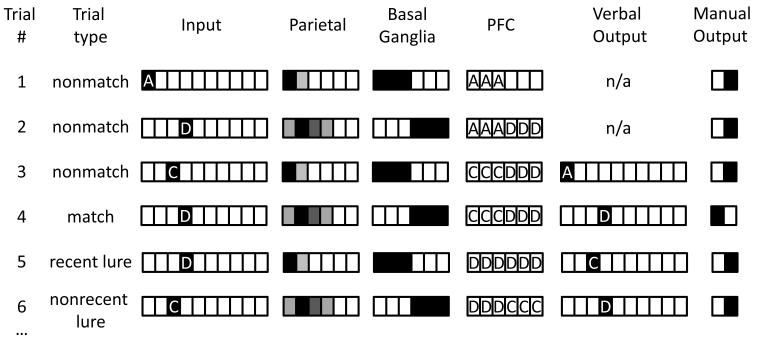
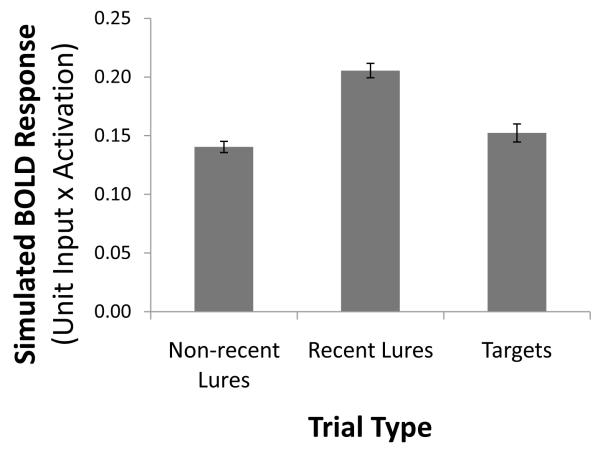
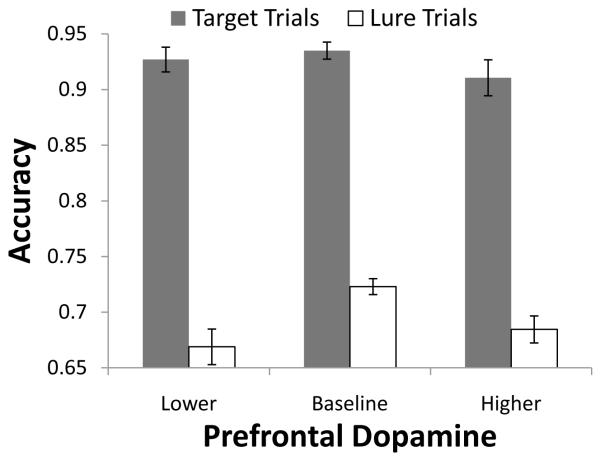
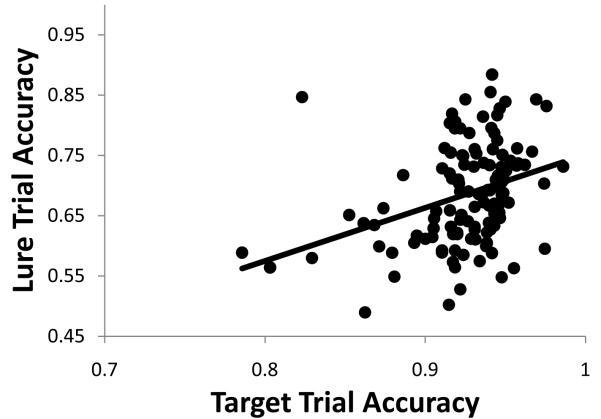
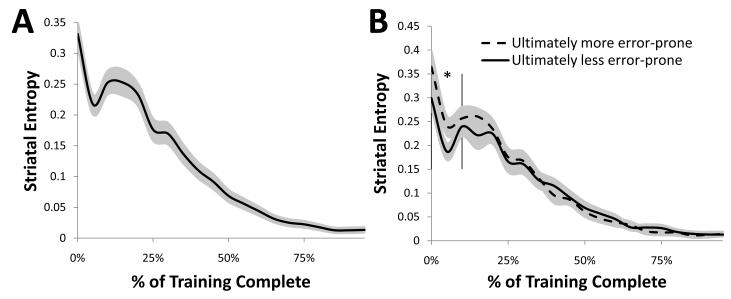
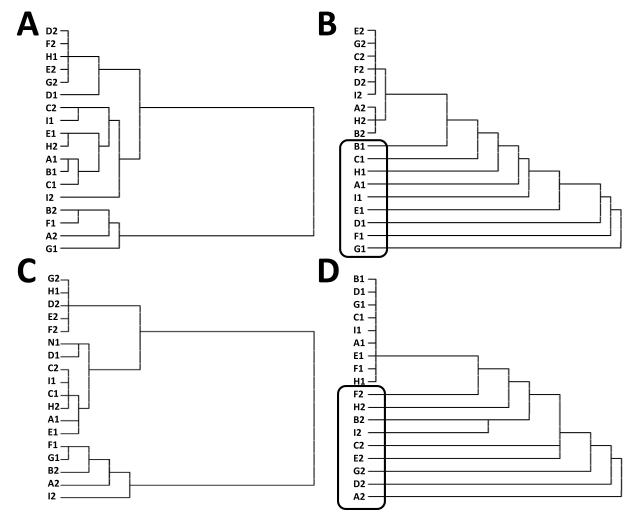
A paradigmatic test of executive control, the n-back task, is known to recruit a widely distributed parietal, frontal, and striatal "executive network," and is thought to require an equally wide array of executive functions. The mapping of functions onto substrates in such a complex task presents a significant challenge to any theoretical framework for executive control. To address this challenge, we developed a biologically constrained model of the n-back task that emergently develops the ability to appropriately gate, bind, and maintain information in working memory in the course of learning to perform the task. Furthermore, the model is sensitive to proactive interference in ways that match findings from neuroimaging and shows a U-shaped performance curve after manipulation of prefrontal dopaminergic mechanisms similar to that observed in studies of genetic polymorphisms and pharmacological manipulations. Our model represents a formal computational link between anatomical, functional neuroimaging, genetic, behavioral, and theoretical levels of analysis in the study of executive control. In addition, the model specifies one way in which the pFC, BG, parietal, and sensory cortices may learn to cooperate and give rise to executive control.
Figures

References
-
- Aalto S, Brück A, Laine M, Någren K, Rinne JO. Frontal and temporal dopamine release during working memory and attention tasks in healthy humans: A positron emission tomography study using the high-affinity dopamine D2 receptor ligand [11C]FLB 457. Journal of Neuroscience. 2005;25:2471–2477. - PMC - PubMed
-
- Apud JA, Weinberger DR. Treatment of cognitive deficits associated with schizophrenia: Potential role of catechol-O-methyltransferase inhibitors. CNS Drugs. 2007;21:535–557. - PubMed
-
- Bilder RM, Volavka J, Lachman HM, Grace AA. The catechol-O-methyltransferase polymorphism: Relations to the tonic-phasic dopamine hypothesis and neuropsychiatric phenotypes. Neuropsychopharmacology. 2004;29:1943–1961. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous