

KIF4 regulates midzone length during cytokinesis
- PMID: 21565503
- PMCID: PMC3100440
- DOI: 10.1016/j.cub.2011.04.019
KIF4 regulates midzone length during cytokinesis
Abstract
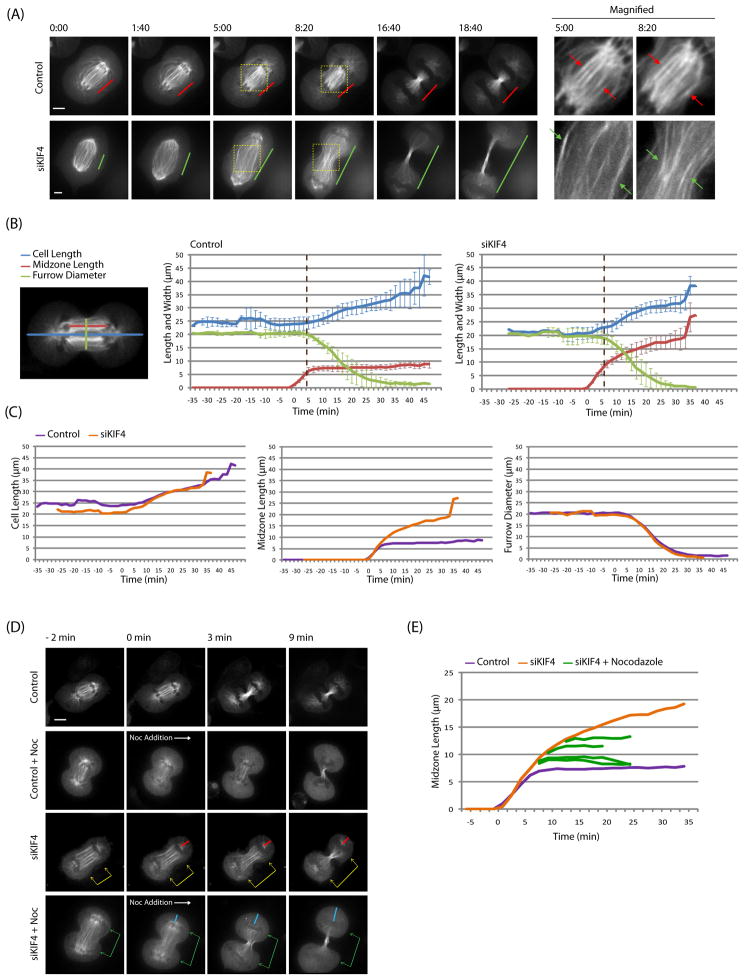
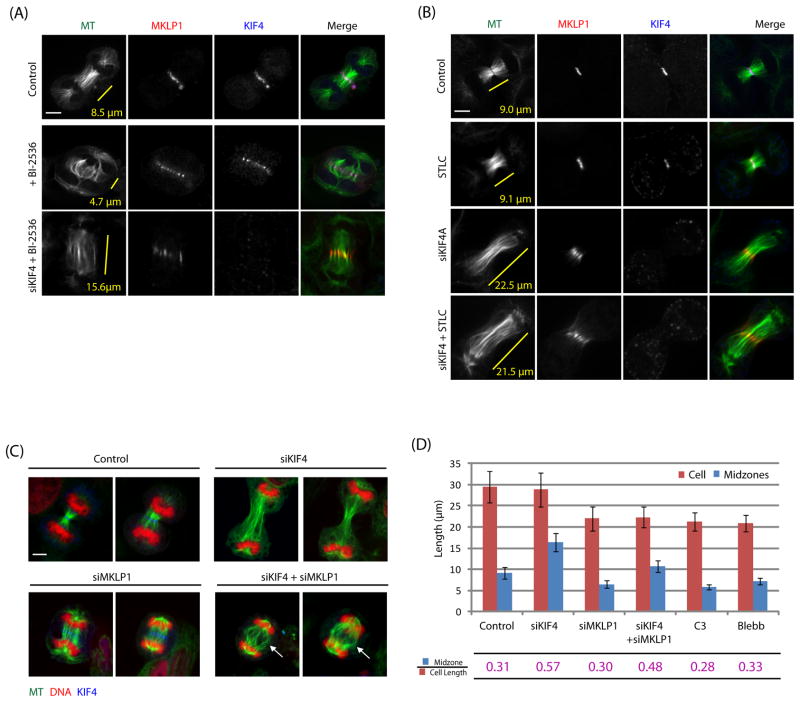
Background: Midzones, also called central spindles, are an array of antiparallel microtubules that form during cytokinesis between the separated chromosomes. Midzones can be considered to be platforms that recruit specific proteins and orchestrate cytokinetic events, such as sister nuclei being kept apart, furrow ingression, and abscission. Despite this important role, many aspects of midzone biology remain unknown, including the dynamic organization of midzone microtubules. Investigating midzone microtubule dynamics has been difficult in part because their plus ends are interdigitated and buried in a dense matrix, making them difficult to observe.
Result: We employed monopolar cytokinesis to reveal that midzone plus ends appear to be nondynamic. We identified the chromokinesin KIF4 as a negative regulator of midzone plus-end dynamics whose activity controls midzone length but not stability. KIF4 is required to terminate midzone elongation in late anaphase. In the absence of KIF4, midzones elongate abnormally, and their overlap regions are unfocused. Electron-dense material and midbodies are both absent from the elongated midzones, and actin filaments from the furrow cortex are not disassembled after ingression.
Conclusion: KIF4-mediated midzone length regulation appears to occur by terminating midzone elongation at a specific time during cytokinesis, making midzones and mitotic spindles differ in their dynamics and length-regulating mechanisms.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures




References
-
- Bringmann H. Cytokinesis and the spindle midzone. Cell Cycle. 2005;4:1709–1712. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
